
-
1 植物分类系统
-
2 PPT
一 植物分类系统
对植物的分类可以分为人为分类系统和自然分类系统。人为分类系统是根据人们的使用习惯或经济用途等的分类,与植物之间的亲缘关系没有任何关系。自然分类系统是根据植物的形态特征、亲缘关系、代谢产物等进行的分类。植物分类系统应便于识别、查询和应用植物种类。
园林中使用的植物分类系统多为自然分类系统,由于依据不同,不同分类系统间有差异,常见的分类系统有:恩格勒系统、哈钦松系统、塔赫他间系统和柯朗奎斯特系统。
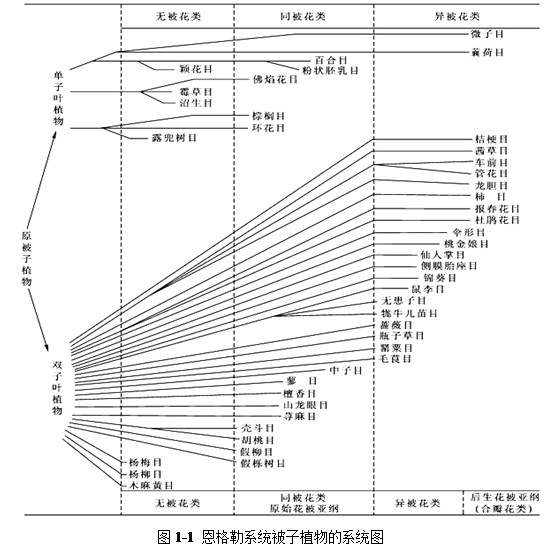
1 恩格勒植物分类系统
由德国植物分类学家恩格勒(A.Engler)和勃兰特(K.Pranti)于1897年在其《植物自然分科志》巨著中所使用的系统,它是分类学史上第一个比较完整的系统,它将植物界分13门,第13门为种子植物门,再分为裸子植物和被子植物两个亚门,被子植物亚门包括单子叶植物和双子叶植物两个纲,并将双子叶植物纲分为离瓣花亚纲(古生花被亚纲)和合瓣花亚纲(后生花被亚纲)。
恩格勒系统将单子叶植物放在双子叶植物之前,将合瓣花植物归并一类,认为是进化的一群植物,将柔荑花序植物作为双子叶植物中最原始的类群,而把木兰目、毛茛目等认为是较为进化的类群,把豆目归为蔷薇目下的一个科等等,这些观点为现代许多分类学家所不赞同(系统图如图1-1)。
恩格勒系统几经修订,在1964年出版的《植物分科志要》第十二版中,已把双子叶植物放在单子叶植物之前。共有62目,344科,其中双子叶植物48目,290科,单子叶植物14目,54科。这个系统在世界各国影响极大。在我国,多数植物研究机关、大学生物系标本馆和出版的分类学著作中被子植物分科多按恩格勒系统第11版排列,如中国植物志、秦岭植物志、内蒙古植物志、河北植物志、北京植物志等。
总的来看,该系统有如下的特点:
(1) 认为单性而无花被是较原始的特征;
(2) 认为单子叶植物较双子叶植物为原始;
(3) 目与科的范围较大。
2 哈钦松植物分类系统
英国植物学家哈钦松(J.Hutchinson)于1926年和1934年在其《有花植物科志》I、 II中所建立的系统。在1973年修订的第三版中,共有111目,411科,其中双子叶植物82目,342科,单子叶植物29目,69科(系统图如图1-2)。
该分类系统的主要特点有:
(1) 认为两性花比单性花原始,花部分离,多数,螺旋状排列的比花各部合生、定数、轮生的进化,虫媒比风媒原始。在现代被子植物中,多心皮类包括木兰目和毛茛目是最原始的;
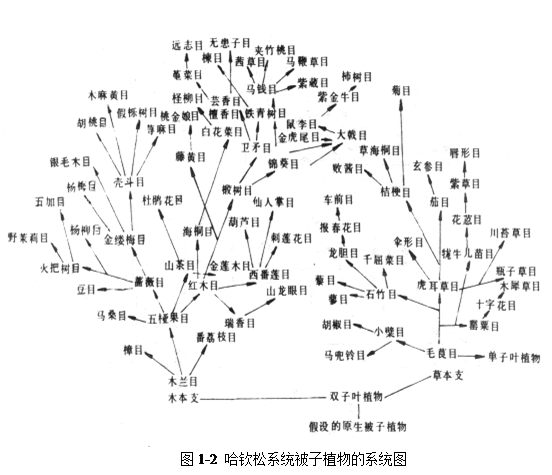
(2)单被花和无被花是次生的,来源于双被花类;柔荑花序类群较进化,起源于金缕梅目;
(3) 单子叶植物和双子叶植物有共同的起源,木本植物起源于木兰目,草本植物起源于毛茛目;
(4) 认为单子叶植物比较进化,故排在双子叶植物之后;
(5) 单叶和叶呈互生排列现象属于原始性状;
(6) 将木本植物与草本植物分开,并认为乔木为原始性状,草本为进化性状;
(7) 分科比较小,较易运用和掌握。
目前在我国,建立较晚的标本室,如中国科学科院昆明植物所、中国科学院华南植物所、中国科学院广西植物所、福建、贵州的经济植物标本室等多用哈钦松系统。南方的高等院校植物标本室也多采用哈钦松系统排列标本。
有人认为1973年版比原版更不好用,比如有些双子叶植物科本来关系较接近,如唇形目与马鞭草目用草本支、木本支为标准在系统树很早被分开,但实际上关系很近,五加科与伞形科亦是如此。人们宁可用旧版而不用它的新版系统,认为新版加重二元思想的色彩。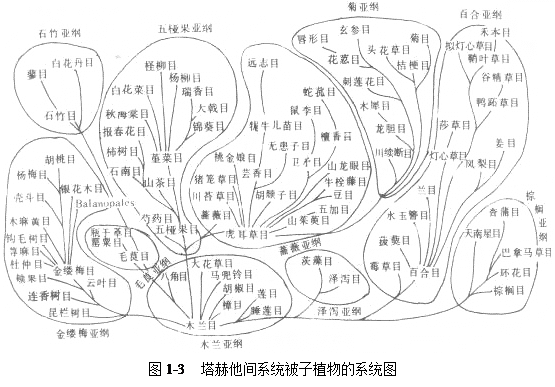
3 塔赫他间植物分类系统
这是前苏联植物学家塔赫他间(A.Takhtajan)于1954年在其《被子植物起源》一书中公布的系统,这一学说是在真花说的基础上建立起来的。该系统打破了传统把双子叶植物分为离瓣花亚纲和合瓣花亚纲的分类;在分类等级上增设了“超目”一级分类单元。将原属毛茛科的芍药属独立成芍药科等,这些都和当今植物解剖学、孢粉学、植物细胞分类学和化学分类学的发展相吻合,在国际上得到共识。
塔赫他间系统经过多次修订,在1980年修订版中,共有28超目,92目,416科,其中双子叶植物(木兰纲)20超目,71目333科,单子叶植物(百合纲)8超目,21目,77科,显得较繁锁(系统图如图1-3)。
该系统具有以下特点:
(1) 被子植物起源于拟苏铁类;
(2) 由木本植物演化出草本植物,因此木本植物为原始的类型;
(3) 单子叶植物起源于水生双子叶植物类的睡莲目中的莼菜科
(4) 他主张单元起源。由木兰目演化出毛茛目及睡莲目,草本单子叶植物起源于睡莲目;木本单子叶植物则木兰目直接演化而来。柔荑花序类各目起源于金缕梅目。
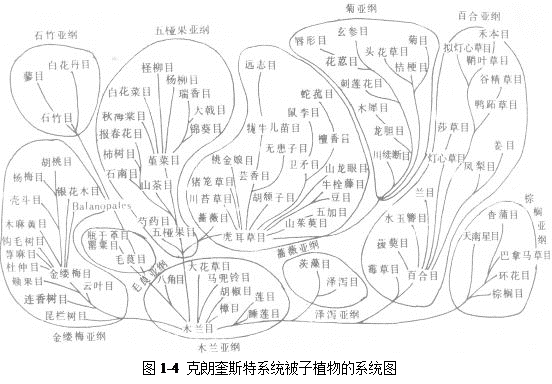
4 克朗奎斯特植物分类系统
克朗奎斯特分类法是由美国学者阿瑟·克朗奎斯特最早于1958年发表的一种对有花植物进行分类的体系,1981年在他的著作《有花植物的综合分类系统》中最终完善。包括64个目和383个科,16.5万种植物。木兰纲(即双子叶植物):包含6亚纲,64目,318科。即木兰亚纲、石竹亚纲、金缕梅亚纲、蔷薇亚纲、五桠果亚纲、菊亚纲。百合纲(即单子叶植物纲):包括5亚纲,19目,65科约6万种植物。即泽泻亚纲、槟榔亚纲、鸭跖草亚纲、姜亚纲、百合亚纲(系统图如图1-4)。这个系统近年在我国有较大影响,各级分类系统的安排上较为合理,科的数目及范围较适中,有利于教学使用。近年来高校一些植物学教科书中,被子植物的讲授多数采用该系统。
该分类系统亦采用真花学说及单元起源的观点,认为有花植物起源一类已经绝灭的种子蕨;木兰目是被子植物的原始类型;柔荑花序类各目起源于金缕梅目。
本书的被子植物采用了此系统。裸子植物采用了邓万钧等的分类系统。
5 被子植物APG分类法
《被子植物APG分类法》是1998年由被子植物种系发生学组(APG)出版的一种对于被子植物的现代分类法。这种分类法和传统的依照形态分类不同,是主要依照植物的三个基因组DNA的顺序,以亲缘分支的方法分类,包括两个叶绿体和一个核糖体的基因编码。虽然主要依据分子生物学的数据,但是也参照其他方面的理论,例如将真双子叶植物分支和其他原来分到双子叶植物纲中的种类区分,也是根据花粉形态学的理论。
APG系统下设分类包括40个目和462个科。有的科分布地域非常狭小或新分出的科,尚未有通用的汉语译名。
二 植物分类的依据
1 按植物形态学进行分类

根据植物器官的外部形态特征,如根、茎、叶、花、果实、种子等,作为分类的重要依据。在形态学分类的依据中,花及果的特征最为分类学家看重,因为它们的性状最为稳定,变异较小。
植物的外部形态非常直观,实践应用过程中也最为容易,因而许多的植物分类检索表多以此为依据。就目前园林中对园林树木的分类而言,基本上是采用形态分类法。如蝶形花科的蝶形花冠,蔷薇科的蔷薇花冠(图1-5)等;豆目三科的荚果,壳斗科的坚果及果苞等。随着放大镜等光学显微镜的使用,很多表皮附属物等的微小形态也成了分类的重要依据,如茸毛、星状毛等。
2 按植物的解剖特征进行分类
利用显微镜对植物体内部组织结构进行观察比较,多用于较高等级分类单元的鉴别。如松属树木针叶的维管束数、树脂道位置及数目为重要的分种特重征。一般情况下,植物解剖学的应用是在形态特征十分相似的情况下进行的。
3 按孢子和花粉进行分类
研究植物的孢子和花粉,并以此作为分类依据。孢粉形态不易受外界环境的影响,是很好的分类证据。特别是扫描电镜的运用,可对花粉壁饰纹形态、沟孔、极性、对称性等进行观察,但是成本较高,往往只是其它分类方法的补充。
4 按细胞的特征进行分类
主要对细胞中染色体数目、结构、形态及行为进行研究,即染色体组型和核型分析。染色体为遗传物质的载体,性状稳定,因而受到分类学家的重视。染色体资料对于验证形态分类结果、分析亲缘关系、识别杂种等具有重要意义。如牡丹属(Paeonia)为毛茛科植物,但其形态与其它种的一般特征有较大区别,后发现其染色体基数x=5,而该科其它属x=6~10,13,据此,有的学者将其从毛茛科中分出成立了牡丹科。
5 根据植物的化学成分进行分类
以植物化学成分依据来解决分类学问题。原因是亲缘关系近的植物类群会有相似的化学成分和产物,故可根据化学成分研究植物类群分类的合理性,分析其亲缘关系。如红豆杉植物含紫杉类生物碱而三尖杉属植物含粗榧碱和刺桐类生物碱,二者化学成分不同,产生途径各异,因而将该属与红豆杉科分开独立成科。
6 根据植物基因组成进行分类
根据植物基因组成的分类其本质是分子分类,广义上的分子分类包括同工酶分析与蛋白质分析。狭义的分子分类是以DNA多态性为基础的分类方法,目前用于揭示DNA多态性的分子标记有限制性片段长度多样性(RFLP)、随机扩增多态性DNA、扩增片段长度多态性(RAPD)、扩增片段长度多态性(AFLP)、简单重复序列(SSR)等。
从理论上讲,直接测定植物DNA序列进行比较,从而进行分类及进化生物学研究是最彻底、最直接的手段,但高等植物DNA极为庞大、难以进行全序列测定,目前某些基因已广泛应用,如叶绿体基因组cpDNA、核基因组nrDNA、线粒体基因组mtDNA等。
植物分类的依据
1 按植物形态学进行分类
根据植物器官的外部形态特征,如根、茎、叶、花、果实、种子等,作为分类的重要依据。在形态学分类的依据中,花及果的特征最为分类学家看重,因为它们的性状最为稳定,变异较小。
植物的外部形态非常直观,实践应用过程中也最为容易,因而许多的植物分类检索表多以此为依据。就目前园林中对园林树木的分类而言,基本上是采用形态分类法。如蝶形花科的蝶形花冠,蔷薇科的蔷薇花冠(图1-5)等;豆目三科的荚果,壳斗科的坚果及果苞等。随着放大镜等光学显微镜的使用,很多表皮附属物等的微小形态也成了分类的重要依据,如茸毛、星状毛等。
2 按植物的解剖特征进行分类
利用显微镜对植物体内部组织结构进行观察比较,多用于较高等级分类单元的鉴别。如松属树木针叶的维管束数、树脂道位置及数目为重要的分种特重征。一般情况下,植物解剖学的应用是在形态特征十分相似的情况下进行的。
3 按孢子和花粉进行分类
研究植物的孢子和花粉,并以此作为分类依据。孢粉形态不易受外界环境的影响,是很好的分类证据。特别是扫描电镜的运用,可对花粉壁饰纹形态、沟孔、极性、对称性等进行观察,但是成本较高,往往只是其它分类方法的补充。
4 按细胞的特征进行分类
主要对细胞中染色体数目、结构、形态及行为进行研究,即染色体组型和核型分析。染色体为遗传物质的载体,性状稳定,因而受到分类学家的重视。染色体资料对于验证形态分类结果、分析亲缘关系、识别杂种等具有重要意义。如牡丹属(Paeonia)为毛茛科植物,但其形态与其它种的一般特征有较大区别,后发现其染色体基数x=5,而该科其它属x=6~10,13,据此,有的学者将其从毛茛科中分出成立了牡丹科。
5 根据植物的化学成分进行分类
以植物化学成分依据来解决分类学问题。原因是亲缘关系近的植物类群会有相似的化学成分和产物,故可根据化学成分研究植物类群分类的合理性,分析其亲缘关系。如红豆杉植物含紫杉类生物碱而三尖杉属植物含粗榧碱和刺桐类生物碱,二者化学成分不同,产生途径各异,因而将该属与红豆杉科分开独立成科。
6 根据植物基因组成进行分类
根据植物基因组成的分类其本质是分子分类,广义上的分子分类包括同工酶分析与蛋白质分析。狭义的分子分类是以DNA多态性为基础的分类方法,目前用于揭示DNA多态性的分子标记有限制性片段长度多样性(RFLP)、随机扩增多态性DNA、扩增片段长度多态性(RAPD)、扩增片段长度多态性(AFLP)、简单重复序列(SSR)等。
从理论上讲,直接测定植物DNA序列进行比较,从而进行分类及进化生物学研究是最彻底、最直接的手段,但高等植物DNA极为庞大、难以进行全序列测定,目前某些基因已广泛应用,如叶绿体基因组cpDNA、核基因组nrDNA、线粒体基因组mtDNA等。

