影响群落组成和结构的因素
1.生物因素
群落结构在其形成过程中,生物因素起着重要的作用,其中作用最大的是竞争和捕食。
1)竞争对群落结构的影响
竞争是生物群落结构形成的一个重要驱动因素。一般认为,生物群落中物种通过竞争引起生态位分化,从而增加群落的物种多样性。MacArthur曾研究了北美针叶林中林莺属(Dendroica)的5种食虫小鸟,发现它们在树上的不同部位取食,形成资源分隔现象,可以被解释为因竞争而产生的共存。人工控制条件下的去除实验表明,在美国亚利桑那荒漠中有一种可格卢鼠和三种囊鼠共存,其栖息的小环境和食性上彼此有所区别,当去除其中某一种时,另外三种的小生境就有明显的扩大。这些实例都反证了竞争对群落结构的影响。
2)捕食对群落结构的影响
捕食对群落结构形成的影响,视捕食者是泛化种还是特化种而异。如果捕食者是泛化种,随着捕食者食草压力的增加,可使草地的植物多样性增加,而当捕食压力过高时,又会导致植物多样性下降。如果捕食者是特化种,当其捕食对象为群落的优势种时,可导致群落多样性的增加,而当捕食对象为竞争力弱的种时,随着捕食压力的增加,物种多样性就会呈线性下降趋势。
2.干扰对群落结构的影响
干扰(disturbance)是自然界的普遍现象,生物群落不断经受各种随机变化的事件的干扰,引起群落的非平衡特性,对群落的结构形成和存在状态有重要影响。
1)中度干扰假说
Cornell等提出中等程度的干扰频率能维持群落较高多样性的假说(intermediate disturbance hypothesis)。其理由是:①一次干扰后少数先锋物种入侵断层,如果干扰频繁、一再出现的话,则先锋物种不能发展到演替的中期,物种多样性保持较低;②如果干扰间隔期很长,使演替能发展到顶极期,竞争排斥起到了排斥他种的作用,多样性也不高;③只有中等程度的干扰将使多样性最高,它允许更多的物种入侵和建立种群。
2)干扰与群落的缺口
连续的群落出现缺口(gaps)是非常普遍的现象,缺口的形成往往是由于干扰而造成的。例如大风、雷电、火烧、雪崩、砍伐等引起森林群落的缺口,而冻融、动物挖掘、啃食、践踏等可引起草地群落的缺口。干扰造成群落缺口后,有的在没有继续干扰的条件下会逐渐按照当地的典型演替序列而出现可以预测的有序演替过程,但也有的则经受完全不可预测的变化,缺口可能被周围群落中的任何一物种入侵和占据,并发展为优势种。
3.空间异质性和群落结构
环境条件不是均匀一致的,从而导致群落的空间异质性(spatial heterogeneity)。空间异质性越高,群落的小生境越多,群落会有更多的物种存在。
研究表明,在土壤和地形变化频繁的地段,植物群落含有更多的物种,而平坦同质土壤上群落的物种多样性偏低。淡水系统中,底质类型越多,软体动物种类越多。MacArthur等研究鸟类多样性与植物物种多样性和取食高度多样性之间的关系,发现鸟类多样性与植物多样性的相关不如与取食高度多样性的相关明显,说明对于鸟类生活来说,植被的分层结构比物种组成更重要。在灌丛和草地群落中,垂直分层不如森林明显,而水平结构的异质性就起到决定性作用。
4.岛屿与群落结构
岛屿的物种数与面积的关系 通常岛屿上(或一个地区中)物种数目会随着岛屿面积的增加而增加,最初增加十分迅速,当物种接近该生境所能承受的最大数量时,增加将逐渐停止。
海岛的物种数-面积关系,可用下述方程描述:S=cAz
或取对数 lgS=lgC + Z(lgA)
其中:S=种数,A=面积,Z和C为两个常数,Z表示物种数-面积关系中回归的斜率,
C是表示单位面积物种数的常数。
岛屿面积越大种数越多,称为岛屿效应,因为岛屿处于隔离状态,其迁入和迁出的强度低于周围连续的大陆。Lack认为,大陆具有较多物种数是含有较多的生境的简单反映,即生境多样性导致物种多样性。
MacArthur的平衡说 岛屿上的物种数决定于物种迁入和灭亡的平衡 。
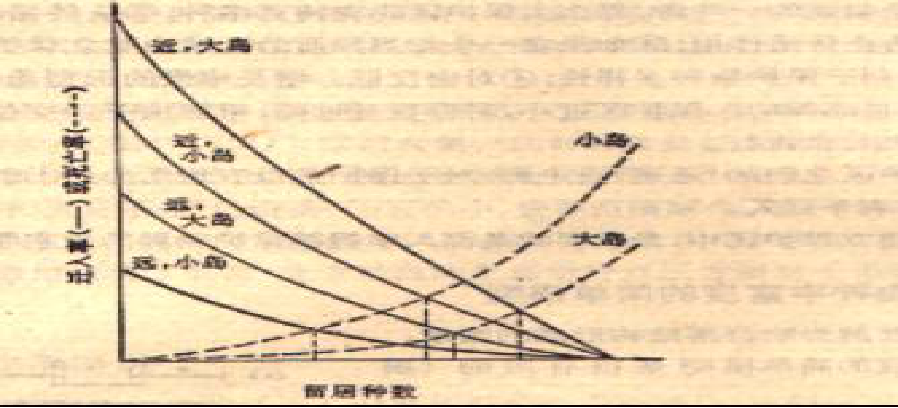
岛屿面积越大且距离大陆越近的岛屿,其留居物种的数目最多,而岛屿面积越小且距离大陆越远的岛屿,其留居物种的数目最少。
根据平衡说,可预测下列四点 :
岛屿上的物种数不随时间而变化
这是一种动态平衡,即灭亡种不断地被新迁入的种所替代
大岛比小岛能“供养”更多的种
随岛距大陆的距离由近到远,平衡点的种数不断降低
岛屿群落的进化
岛屿的物种进化较迁入快,而在大陆,迁入较进化快。
离大陆遥远的岛屿上,特有种可能比较多,尤其是扩散能力弱的分类单元更有可能。
岛屿群落有可能是物种未饱和的,其原因可能是进化的历史较短,不足以发展到群落饱和的阶段。
5 一个物种丰富度的简单模型
物种丰富度的模型可以帮助我们理解影响群落结构形成的因素 。
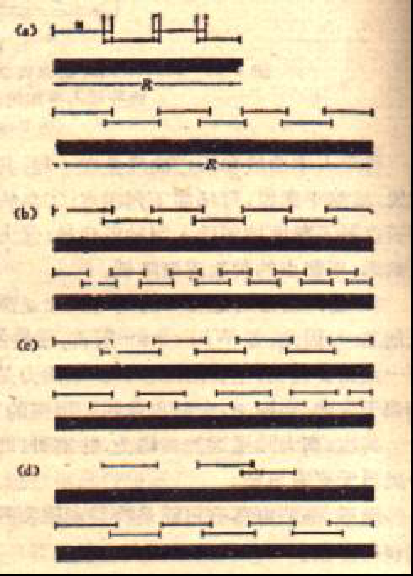
设n为生态位平均宽度,o为生态位重叠度R为群落的有效资源范围。
当n 和o一定时,那么R值越大(代表资源范围大),群落将含有更多的种数。
当R一定时,那么n越小(表示种在利用资源上越分化,生态位越狭),群落中将有更高的物种丰富度
当R一定时,那么o越大(表示物种间利用资源中重叠利用多),群落将含有更多的种数。
当R是一定时,群落的饱和度越高,就越能含有更多的物种数;相反,群落中有一部分资源未被利用,所含种数也就越少。
6.平衡学说与非平衡学说
对于形成群落结构的理论,存在着平衡学说(equilibrium theory)和非平衡学说(non-equilibrium theory)两种对立观点。
平衡学说把生物群落视为存在于不断变化的物理环境中的稳定实体,生活在同一群落中的物种种群处于一种稳定状态。其中心思想是:①群落中共同生活的物种通过竞争、捕食和互利共生等种间作用而相互牵制;②生物群落具有全局稳定性的特点,种间相互作用导致群落的稳定特性,在稳定状态下群落的组成和各物种的数量都变化不大;③群落出现变化实际上是由于环境的变化,即所谓的干扰造成的,并且干扰是逐渐衰亡的。
非平衡学说的主要依据是中度干扰理论。该学说认为,构成群落的物种始终处于变化之中,群落不能达到平衡状态,自然界的群落不存在全局稳定性,有的只是群落的抵抗力(群落抵抗外界干扰的能力)和恢复力(群落受到干扰后恢复到原来状态的能力)。
平衡学说和非平衡学说除对干扰的作用强调不同以外,基本的区别在于:平衡学说的注意点是群落处于平衡点的性质,而对于时间和变异性注意不够;非平衡学说则把注意力放在离开平衡点是群落的行为变化过程,特别强调了时间和变异性。


