1 群落的结构要素
群落的结构包括群落的物理结构和群落的生物结构两个方面。群落的物理结构是指外貌和生长型、垂直分层结构、群落外貌的昼夜和季相。群落的生物结构是指群落的物种组成、种间关系、多样性和演替等。群落的生物结构取决于物理结构。这里主要介绍群落的物理结构。
1.生活型
生活型(life form)是生物对外界环境适应的外部表现形式。关于生活型的划分,早期人们习惯根据植物的形状、大小、分支等外貌特征,同时考虑到植物的生命期长短,把植物分为乔木、灌木、藤本植物、附生植物和草本植物等。目前广泛采用的是丹麦的植物学家C.Raunkiaer系统,他按休眠芽或复苏芽所处的位置高低和保护方式,把高等植物划分为五个生活型,在各类群之下,根据植物体的高度、芽有无芽鳞保护、落叶或常绿、茎的特点等特征,再细分为若干较小的类型(图4-3)。
(1)高位芽植物(phanerophyte):休眠芽或顶端嫩枝位于离地面25cm以上的枝条上,如乔木、灌木等。其中根据体型的高矮又可分为大高位芽植物(芽高度>30m)、中高位芽植物(高度8~30m)、小高位芽植物(高度2~8m)和矮高位芽植物(高度25cm~2m)等类型。
(2)地上芽植物(chamaephyte):植物的芽或顶端嫩枝位于地表土壤表面之上、25cm之下,受土表或残落物所保护,多为灌木、半灌木或草本植物。
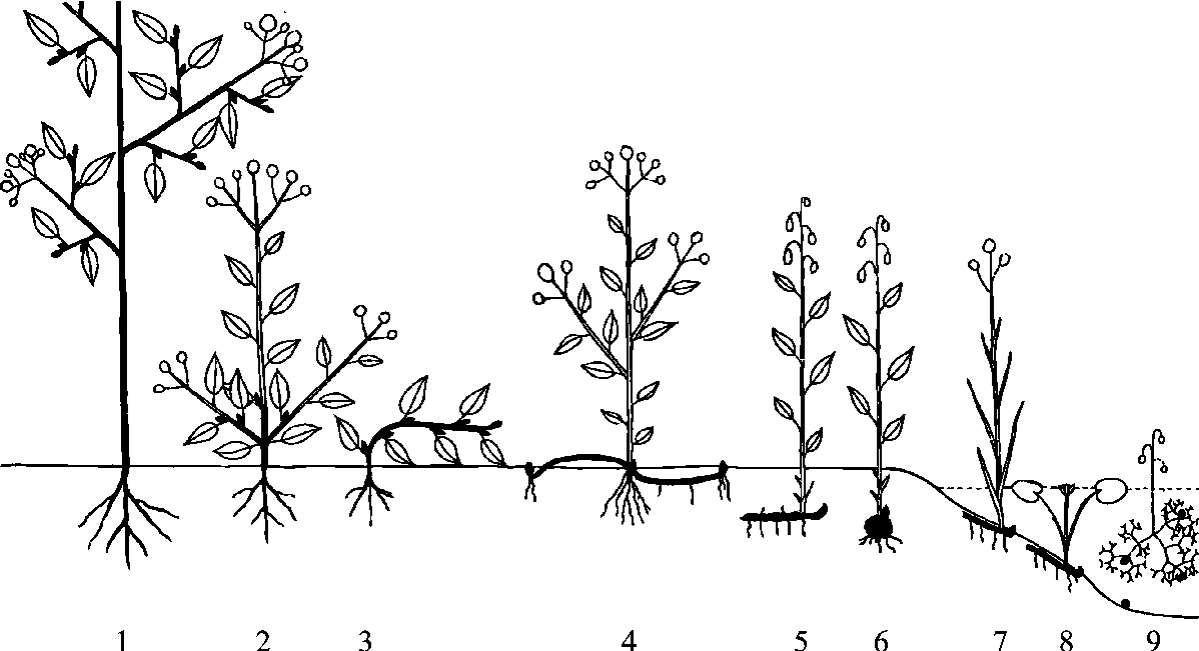
图4-3 Raunkiaer生活型图解
(C.Raunkiaer,1934)
1—位芽植物;2、3—地上芽植物;4—地面芽植物;5~9—隐芽植物

(3)地面芽植物(hemicryptophyte):植物在不利季节,其地上部分死亡,但被土壤和残落物保护的地下部分仍活着,更新芽位于地面土层内。为多年生草本植物。
(4)隐芽植物(cryptophyte):或称地下芽植物(geophyte),植物芽位于较深土层中,或位于水中,多为鳞茎类、块茎类或根茎类多年生草本植物或水生植物。
(5)一年生植物(therophyte):植物只能在良好的季节中生长,它们以种子的形式度过不良季节。
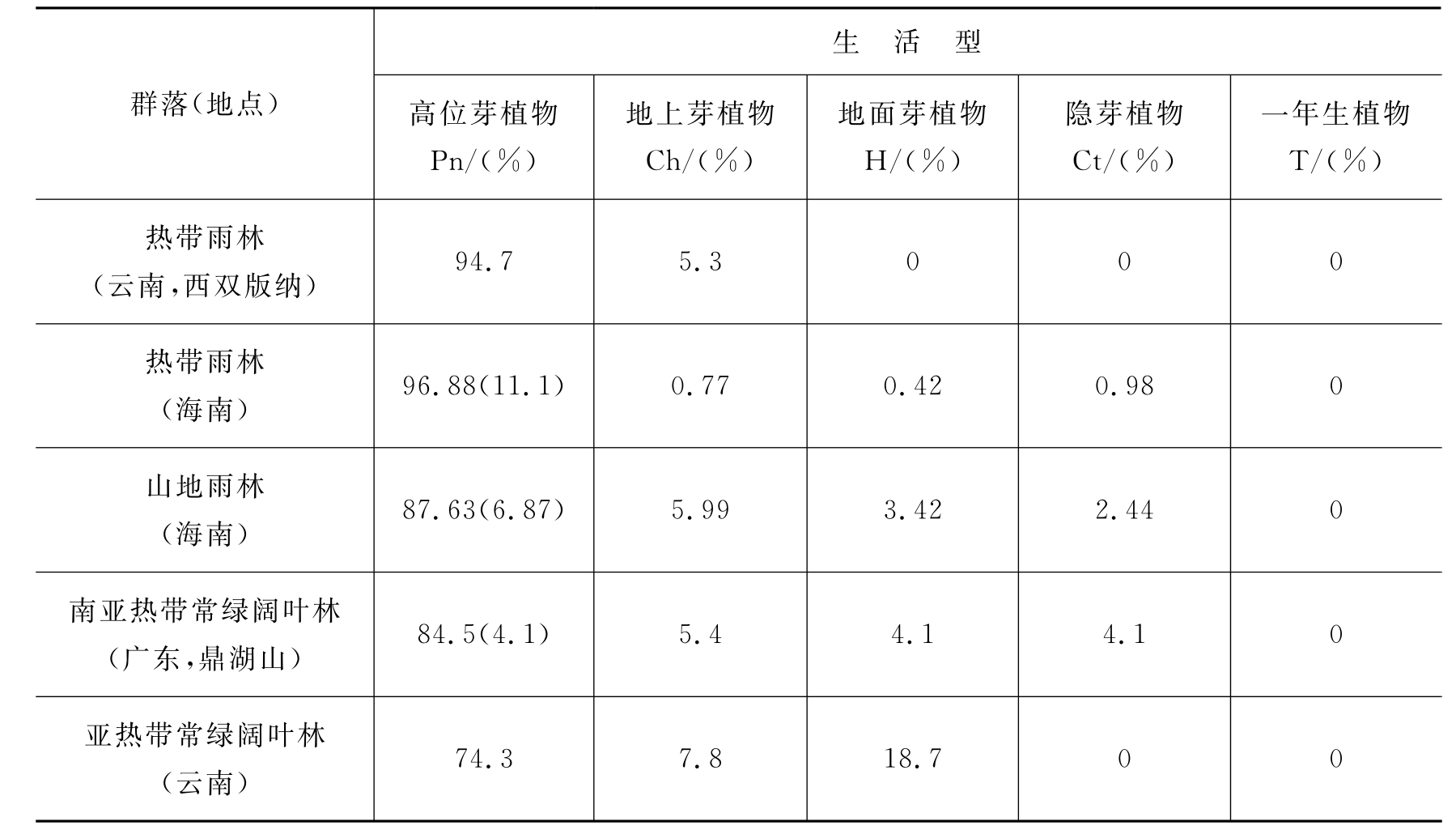
统计各个群落内的各种生活型的数量对比关系称为生活型谱。群落类型不同,其生活型谱也不同。我国自然条件复杂,不同气候区域的主要群落类型中生活型的组成各有特点(表4-2)。
表4-2 我国几种群落类型的生活型谱
续表
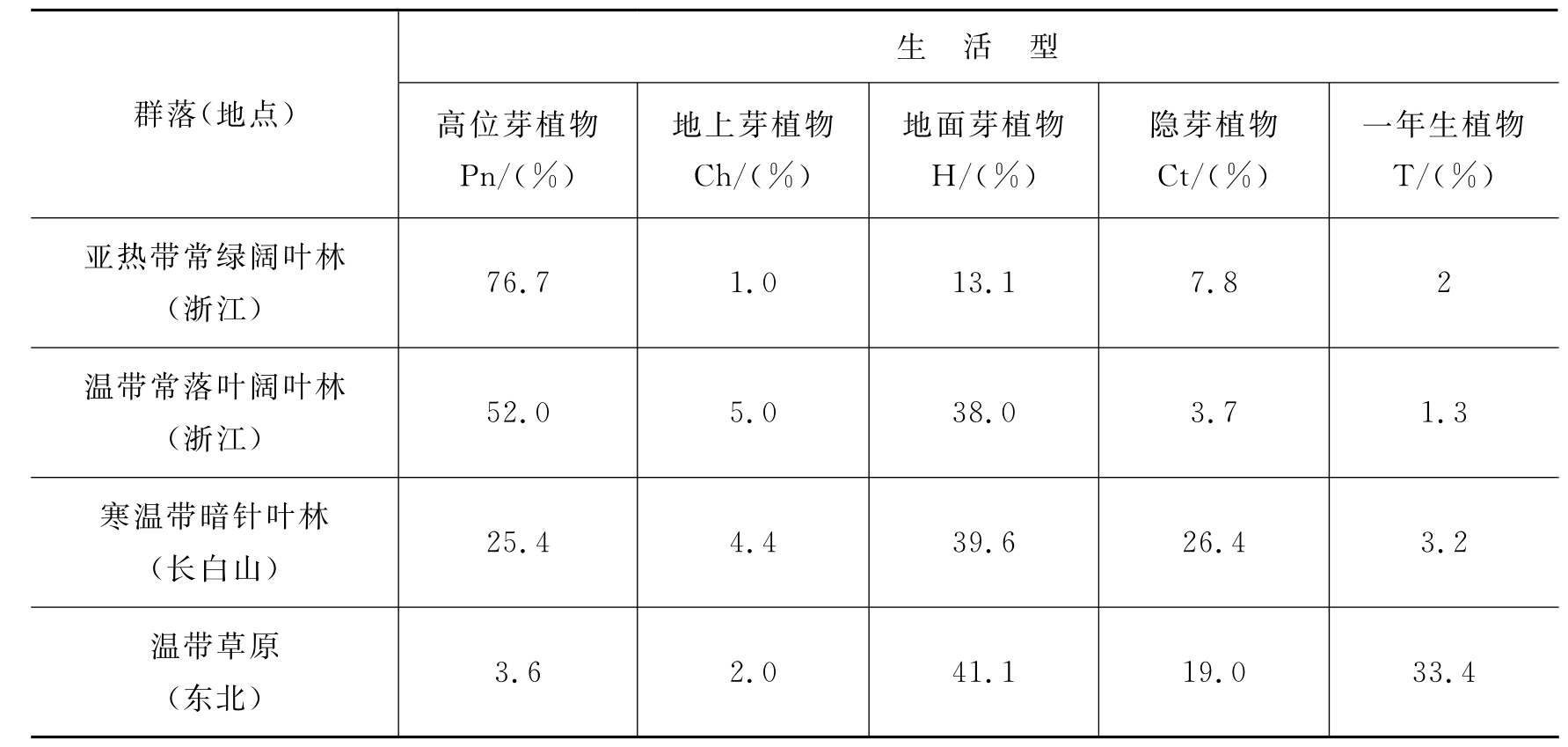
注:引自王伯荪,1987;括号内的数字是指其中藤本植物的百分数。
从表4-2可见,每一类植物群落都是由几种生活型植物所组成的,但其中有一类生活型占优势。这种生活型与环境关系密切:高芽位植物占优势是温暖多湿气候地区群落的特征,如热带雨林群落;地面芽植物占优势,反映了该地区具有较长的严寒季节,如寒温带针叶林群落;地上芽植物占优势,反映了该地区环境比较冷湿;一年生植物占优势则是干旱气候的荒漠和草原地区群落的特征,如温带草原群落。
动物也有不同的生活型,例如兽类中有空中飞行的(蝙蝠)、滑翔的(鼯鼠)、游泳的(鲸、海豹)、地下穴居的(啮齿动物)、奔跑的(马、鹿)等,它们各有各的形态、生理和行为特征,适应各种不同的生活方式。但动物的生活型不能决定生物群落的外貌和结构。
2.叶片性质与叶面积指数
1)叶片大小及性质
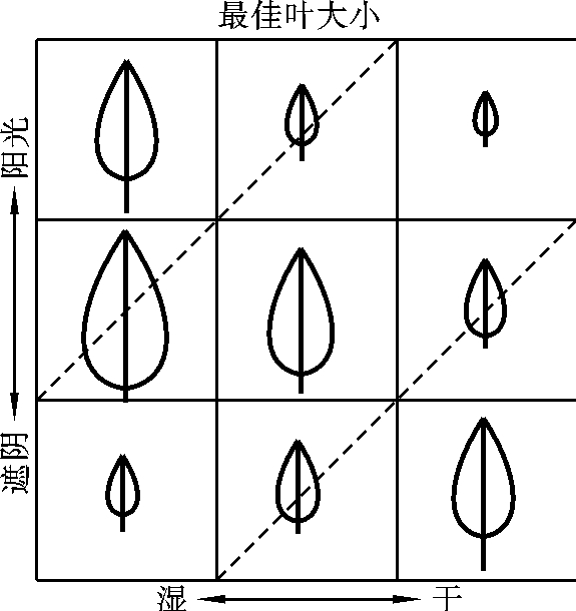
图4-4 最佳叶片大小模型预测的叶片大小与光辐射和水分的关系
(C.J.Krebs,1985)
植物叶片是进行光合作用的主要器官,其大小、形状和性质直接影响着群落的结构与功能。叶的性质如阔叶、针叶、常绿、落叶等也是决定群落外貌的重要特征。叶片的大小与水分平衡和光合收益的效果有密切的关系,而叶温又影响着光合速率。在阳光辐射条件下,大叶比小叶的叶温高,蒸腾量大;相反,在遮阴条件下,大叶的叶温降得快。所以,植物叶片的大小是以平衡蒸腾失水的植物的根吸水量作为成本,以光合收益高低作为收益来确定最佳大小的。C.J.Krebs(1985)根据这种收益-成本分析,预测了在各种光照和土壤水分条件下的最佳叶片大小(图4-4)。
2)叶面积指数
叶面积指数(leaf area index,LAI)是指单位面积土地上单面叶的总面积,是群落结构的一个重要指标,与群落的功能有着直接的关系。
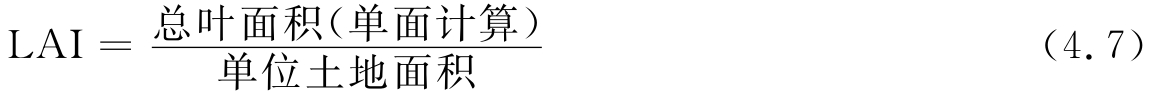
表4-3列出了主要天然植物群落的叶面积指数,可以看出,叶面积指数与群落的光能利用率有直接的关系。
表4-3 主要天然群落类型的叶面积指数与光能利用率

注:引自Barbour,1987;光能利用率为单位面积全年接受的有效光合辐射与该面积上的净生产量之比。
3)层片
层片(synusia)是指群落中由同一生活型的不同植物构成的组合。层片具有以下特征:①属于同一层片的植物是同一生活型类别,但只有当其个体数量相当多,而且相互之间存在一定联系时才能组成层片;②每个层片在群落中均具有一定的小环境,不同层片小环境相互作用构成群落的环境;③每个层片在群落中都占据一定的空间和时间,层片的时空变化形成了群落的不同结构特征。
层片与通常所说的层既有相同之处,又有本质的区别。例如针阔叶混交林的乔木层就包含了针叶和阔叶两个生活型,北方的夏绿阔叶林乔木层可能属同一层片,而热带森林的乔木层可能包含若干个不同的层片。
4)生态位
生态位(niche)是种在群落中的机能作用和地位。根据Г.Ф.Гаузе实验结果,生态位完全相同的种是不能共存的。因此,群落中各个物种是具有一定的生态位分化的,群落越复杂,生态位多样性越高。可以通过计算生态位宽度和生态位重叠,来评价群落中各物种对资源的利用程度和竞争情况。
5)同资源种团
群落中以同一方式利用共同资源的物质集团称为同资源种团(guilds),例如热带地区取食花蜜的许多蜂鸟就是一个同资源种团。
同资源种团是由生态学特征上很相似的种类所组成的,它们彼此之间具有较高的生态位重叠,种间竞争很激烈,一个种因某种原因从群落中消失,其他种就可以取而代之。相比之下,某一同资源种团与群落中其他同资源种团间的关系就较弱。根据同资源种团的特点,可以利用它进行竞争和群落结构的实验研究。同时,以同资源种团作为组成群落的成员,与以物种为组成成员相比,研究会简单得多,这有助于深入研究群落的营养结构。因而研究同资源种团将是群落生态学研究的一个很有希望的方向。
6)群落外貌
群落外貌(physiognomy)是指生物群落的外部形态或表相。它是群落中生物与生物间、生物与环境间相互作用的综合反映。陆地群落外貌主要取决于植被的特征,植物群落是植被的基本单元。水生群落外貌主要取决于水的深度和水流特征。
陆地群落外貌由组成群落的优势生活型和层片结构所决定。群落外貌常随时间的推移而发生周期性变化,这是群落结构的另一重要特征。在一年内随着气候季节变化,群落呈现不同的外貌,这就是季相。
2 群落的垂直结构
1.成层现象
由于环境的逐渐变化,导致对环境有不同需求的动、植物生活在一起,这些动、植物各有其生活型,其生态幅度和适应特点也各有差异,它们各自占据一定的空间,并排列在空间的不同高度和一定土壤深度中。群落这种垂直分化就形成了群落的层次,称为群落垂直成层现象(vertical stratification)。群落的分层现象主要取决于植物的生活型。动物也有分层现象,但不明显。水生环境中,不同的动、植物也在不同深度水层中占有各自位置。
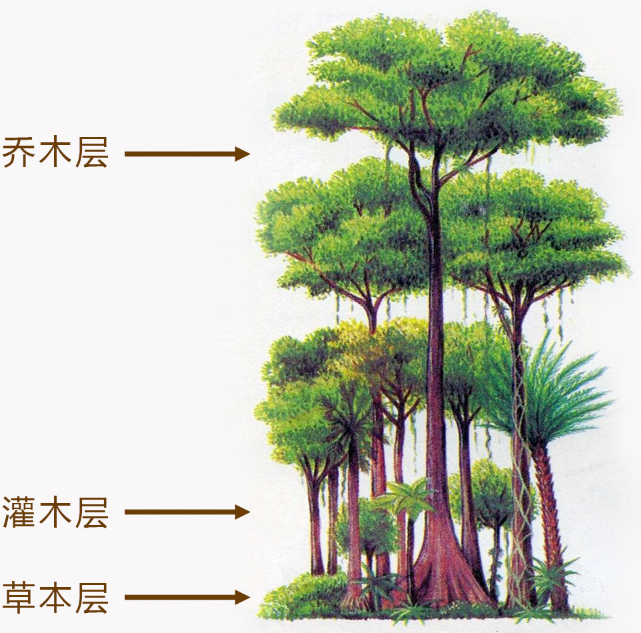
群落的成层现象保证了生物群落在单位空间中更充分地利用自然条件。成层现象发育最好的是森林群落,林中有林冠(canopy)、下木(understory tree)、灌木(shrub)、草本(herb)和地被(ground)等层次。林冠直接接受阳光,是进行初级生产过程的主要地方,其发育状况直接影响到下面各层次。如果林冠是封闭的,林下的灌木和草本植物就发育不好;如果林冠是相当开阔的,林下的灌木和草本植物就发育良好。
以陆生植物为例,成层现象包括地上部分和地下部分。决定地上部分分层的环境因素主要是光照、温度等条件,而决定地下分层的主要因素是土壤的物理化学性质,特别是水分和养分。由此可看出,成层现象是表现植物群落与环境条件相互关系的一种特殊形式。环境条件愈好,群落的层次就愈多,层次结构就愈复杂;环境条件愈差,层次就愈少,层次结构也就愈简单。
2.主要层的作用
多层次结构的群落中,各层次在群落中的地位和作用不同,各层中植物种类的生态习性也是不同的。如以一个郁闭森林群落来说,最高的那一层既是接触外界大气候变化的“作用面”,又因其遮蔽阳光的强烈照射,而保持林内温度和湿度不致有较大幅度的变化。也就是说,这一层在创造群落内的特殊小气候环境中起着主要作用,它是群落的主要层,这一层的树种多数是阳性喜光的种类。上层以下各层次中的植物由上而下耐阴性递增,在群落底层光照最弱的地方则生长着阴性植物,它们不能适应强光照射和温度、湿度的大幅度变化,在不同程度上依赖主要层所创造的环境而生存。由这些植物所构成的层次在创造群落环境中起着次要作用,是群落的次要层,该层中植物的种类常因主要层的结构变化而有较大的变化。区别主要层和次要层,完全按群落中的地位和作用而定。在一般情况下最高的一层通常是主要层,但在特殊情况下,群落中较低的层次也可能是主要层。如热带稀树干草原植被,其分布地气候特别干热,树木星散分布,树冠互不接触,干旱季节全部落叶,在形成植物环境方面作用较小,而密集深厚的草层却强烈影响着土壤的发育,同时也影响着树木的更新。显然,草本层是在群落内占着主要地位的层次。
植物群落中有一些植物,如藤本植物和附、寄生植物,它们并不独立形成层次,而是分别依附于各层次中直立的植物体上,称为层间植物。随着水、热条件愈加丰富,层间植物发育愈加繁茂。粗大木质的藤本植物是热带雨林的特征之一,而附生植物更是多种多样。层间植物主要在热带、亚热带森林中生长发育,而不是普遍生长于所有群落之中,但它们也是群落结构的一部分。
地下(根系)的成层现象和层次之间的关系与地上部分是相应的。一般在森林群落中,草本植物的根系分布在土壤的最浅层,灌木及小树根系分布较深,乔木的根系则深入到地下更深处。地下各层次之间的关系,主要围绕着水分和养分的吸收而实现。
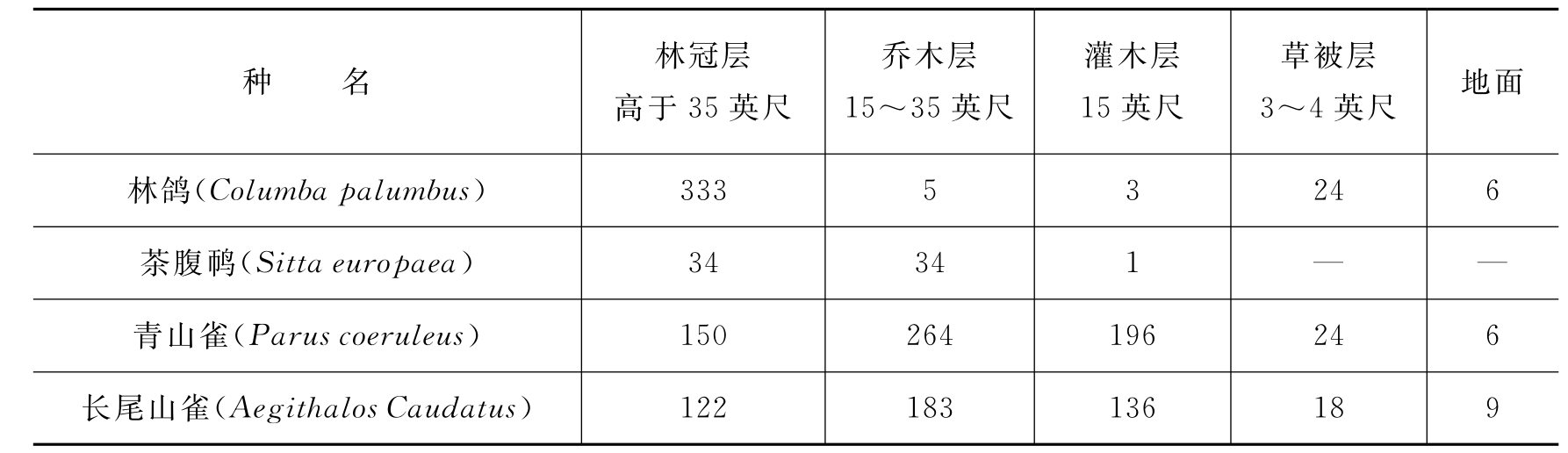
在群落的每一层次中,往往栖息着一些不同程度上可作为各层特征的动物。在群落中动物也有分层现象,如R.H.Whittaker(1952)在美国大烟雾山不同高度的山坡上作了昆虫群落结构分析,有7种昆虫分布在不同垂直高度上,每一物种只局限在一定的高度范围之内(图4-5)。一般说来,群落的垂直分层越多,动物种类也越多。陆地群落中动物种类的多样性,往往是植被层次发育程度的函数。大多数鸟类可同时利用几个不同层次,但每一种鸟却有一个自己所喜好的层次。如表4-4所示,林鸽和茶腹喜欢在林冠层,青山雀、长尾山雀等喜欢在乔木层,沼泽山雀等喜欢在灌木层,而燞鹩等则多在草被层或地面活动。
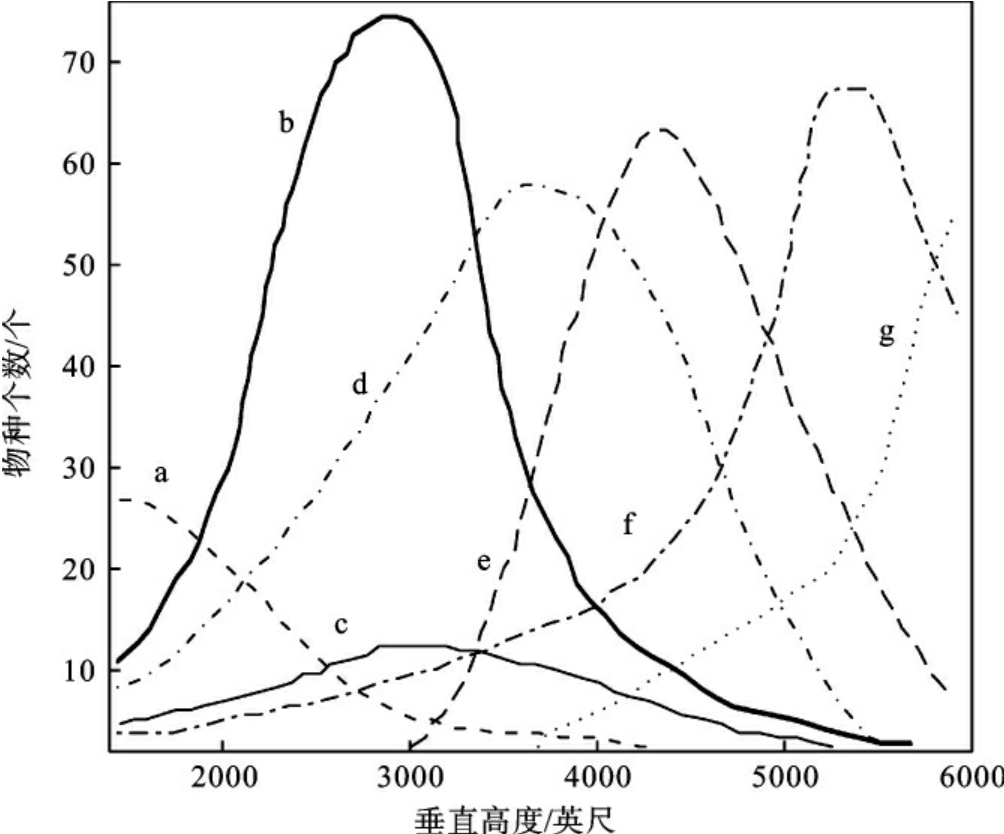
图4-5 美国大烟雾山不同垂直高度上七种昆虫的分布曲线
(仿R.H.Whittaker,1952)
注:其中a、c、f、g是叶蝉,b、d是啮虫,e是芜菁。
表4-4 栎林中鸟类在不同层次中的相对密度
续表
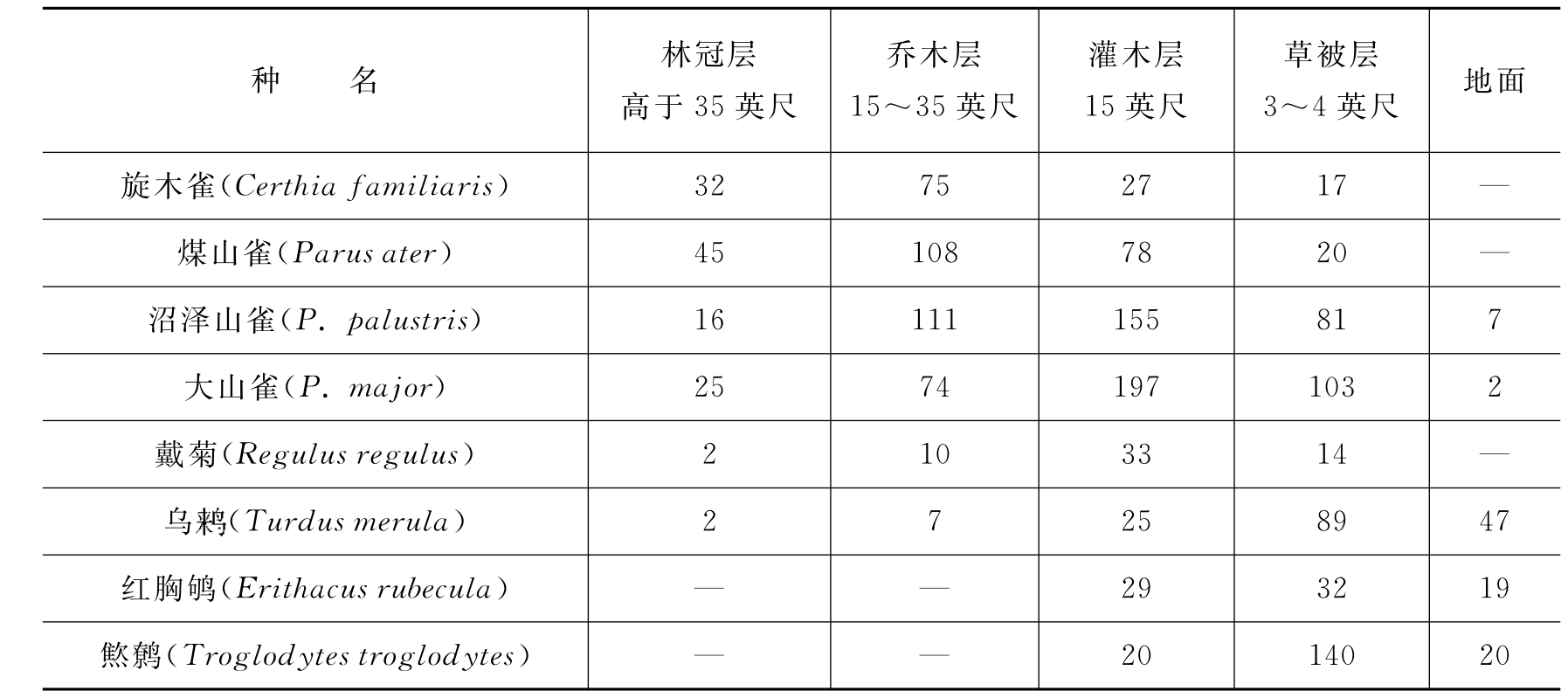
注:1英尺=0.305米。
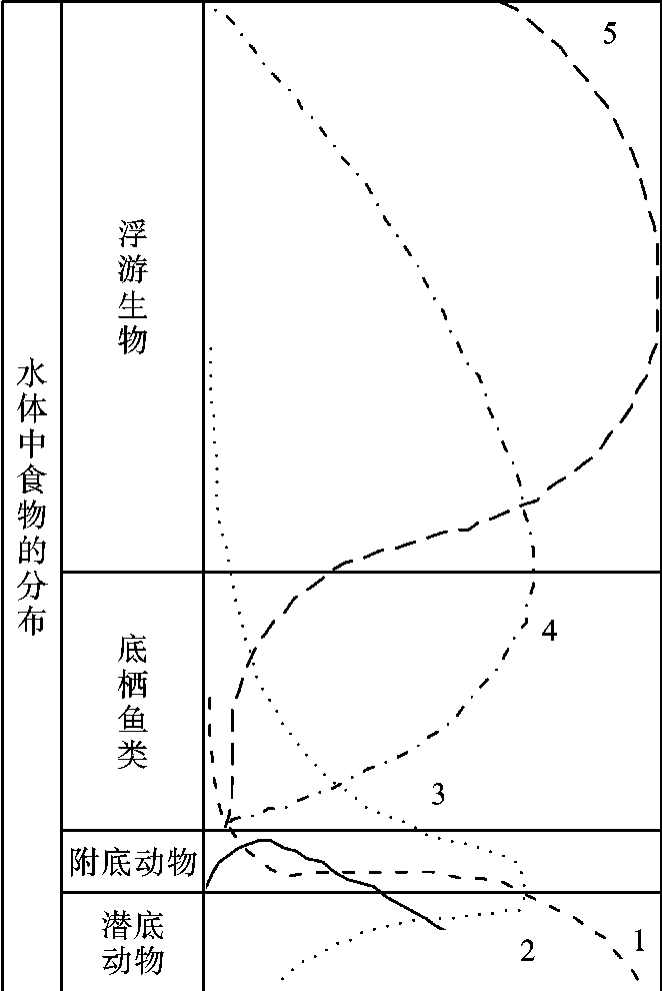
水生群落中,生态位要求不同的各种生物也在不同深度的水体中占据各自的位置,呈现出分层现象。它们的分层主要取决于透光状况、水温和溶解氧的含量。一般可分为漂浮生物(neuston)、浮游生物(plankton)、游泳生物(nekton)、底栖生物(benthos)、附底动物(epifauna)和潜底动物(infauna)等。图4-6是一些鱼类在里海中垂直分布的情况。我国淡水养殖业中的一条传统经验就是在同一水体中混合放养栖息不同水层中的鱼类,以达到提高单产的效果。
3 群落的水平格局
群落结构的另一特征就是水平格局(horizonal pattern),其形成与构成群落的成员的分布状况有关。陆地群落的水平格局主要取决于植物的分布格局。对群落的结构进行观察时,经常可以发现,在一个群落某一地点,植物分布是不均匀的。均匀型分布的植物是少见的。如生长在沙漠中的灌木,由于植株间不可能太靠近,可能比较均匀,但大多数种类是成群型分布。在森林中,林下阴暗的地点,有些植物种类形成小型组合,而林下较明亮的地点是另外一些植物种类形成的组合。在草原中也有同样的情况,在并不形成郁闭植被的草原群落,禾本科密草丛中有与其伴生的少数其他植物,草丛之间的空间,则由各种不同的其他杂草和双子叶杂草所占据。群落内部的这种小型组合可以称为小群落(microcoenosis),它是整个群落的一小部分。小群落形成的原因,主要是环境因素在群落内不同地点上分布不均匀的结果(图4-7),如小地形和微地形的变化、土壤湿度和盐渍化程度的不同,以及群落内植物环境,如上部遮阴不均匀等。同时,植物种类本身的生物学特点也有重大作用,特别是种的繁殖、迁移和竞争等特征,对形成小群落也起到重要作用。
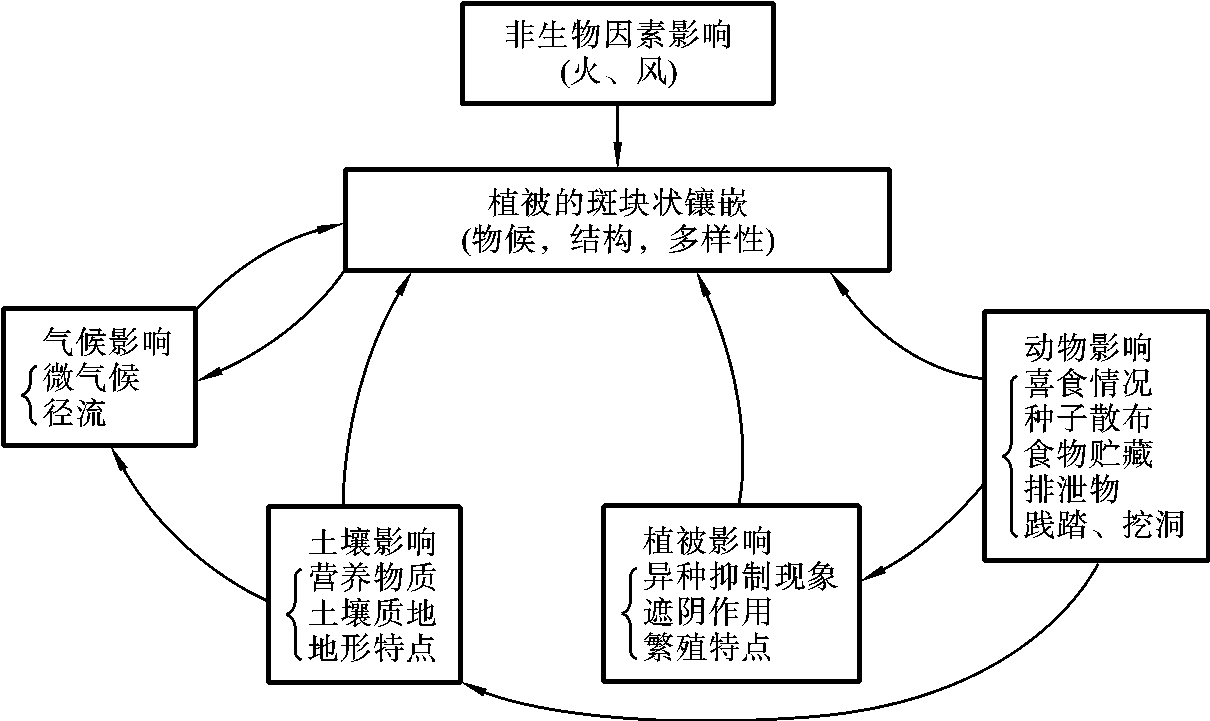
群落水平分化成各个小群落,它们的生产力和外貌特征不相同,在群落内形成不同的斑块。一个群落内出现多个斑块的现象称为群落的镶嵌性(mosaicism),是群落水平分化的一个结构部分。而且,在其形成的过程中依附其所在群落,因此,有人称之为从属群落(subordinate community)。动物群落因其自身的生物学适应范围不同,随着栖息环境的布局而产生相应的水平分布格局。
4 群落的时间格局
很多环境因素具有明显的时间节律,如昼夜节律和季节节律,所以群落结构也随时间而有明显的变化,这就是群落的时间格局(temporal pattern)。相应地,群落中各种植物的生长发育也随之有规律地进行。其中,主要层的植物季节性变化使得群落表现为不同的季节性外貌,即为群落的季相(aspect)。季相变化的主要标志是群落主要层的物候变化。特别是主要层的植物处于营养盛期时,往往对其他植物的生长和整个群落都有着极大的影响,有时当一个层片的季相发生变化时,可影响另一层片的出现与消亡。这种现象在北方的落叶阔叶林内最为显著。早春乔木层片的树木尚未长叶,林内透光度很大,林下出现一个春季开花的草本层片;入夏乔木长叶林冠荫蔽,开花的草本层片逐渐消失。这种随季节而出现的层片,称为季节层片。由于季节不同而出现依次更替的季节层片使得群落结构也发生了季节性变化。群落中由于物候更替所引起的结构变化,又被称为群落在时间上的成层现象。它们在对生境的利用方面起着补充的作用,从而有效地利用了群落的环境空间。
动物群落的季相变化的例子也很多,如人们所熟知的候鸟春季迁徙到北方营巢繁殖,秋季南迁越冬。动物群落的昼夜相也很明显,如森林中,白昼有许多鸟类活动,但一到夜里,鸟类几乎都处于停止活动状态。但一些鸮类开始活动,使群落的昼夜相迥然不同。水生群落的昼夜相不像陆地群落的那么容易看到,但许多淡水和海洋群落中的一些浮游生物有着明显的昼夜相(图4-8)。图中,在日周期中种的个体可上下移动几米,而整个种群在白天则移动到光照最强的水面以下,晚上向上移至水面。阴影多边形表示不同深处的个体相对数。
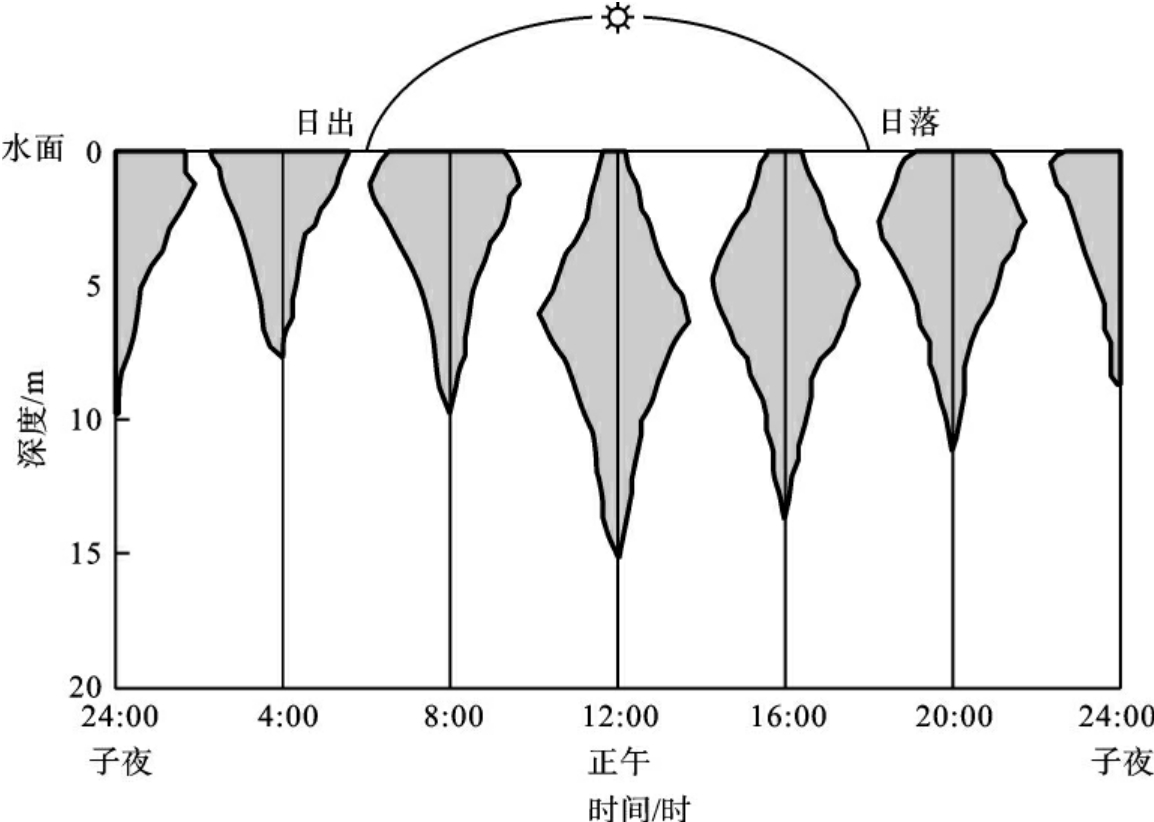
图4-8 一种淡水浮游动物垂直移栖的格局
(仿R.H.Whittaker,1979)
5 群落的交错区和边缘效应
不同群落的交界区域,或两类环境相接触部分,即通常所说的结合部位,称为群落交错区(ecotone)或称生态环境脆弱带。Ecotone为国际生态界最新定义的基本概念之一,一般译作“生态环境交错带”或“生态环境过渡带”。考虑到生态界面的实质,以及该空间域的动态特征,重新将其定义为:在生态系统中,凡处于两种或两种以上的物质体系、能量体系、功能体系之间所形成的“界面”,以及围绕该界面向外延伸的“过渡带”的空间域,称为生态环境脆弱带。它一直被视为界面理论在生态环境中的广延与发展,界面应视为相对均衡要素之间的“突发转换”或“异常空间邻接”。
群落交错区或是生态环境脆弱带实际上是一个过渡地带,这种过渡地带大小不一,有的较窄,有的较宽,有的变化很突然,称为断裂状边缘,有的则表现为逐渐的过渡,或者两种群落互相交错形成镶嵌状,称为镶嵌状边缘(图4-9)。
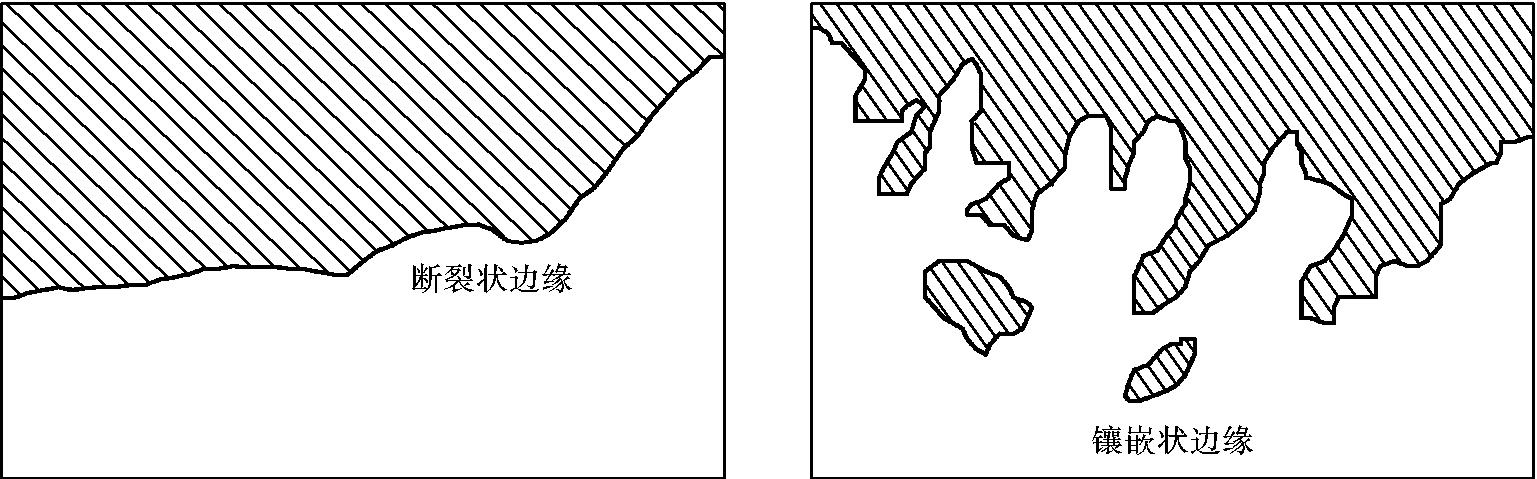
图4-9 两种群落边缘的类型
(仿R.L.Smith,1980)
交错区形成的原因很多,如生物圈内生态系统的不均一,层次结构普遍存在于山区、水域及海陆之间,地形、地质结构与地带性的差异,气候等自然因素变化引起的自然演替、植被分割或景观切割,人类活动造成的隔离,森林、草原遭受破坏,湿地消失和土地沙化等,都是形成交错区的原因。在森林带和草原带的交界地区,常有很宽的森林草原地带,在此地带中,森林和草原镶嵌着出现。群落的边缘有些是持久的,有些是暂时的,这都是环境条件所决定的。
交错区或两个群落的边缘和两个群落的内部核心区域,环境条件往往有明显的区别,如森林草原的边缘,风大、蒸发强,使边缘干燥。太阳辐射在群落的南缘和北缘相差很大,在夏季南向边缘比北向边缘每天可多接受日照数小时,从而使之更加干燥。
在群落交错区内,单位面积内的生物种类和种群密度较之相邻群落有所增加,这种现象称为边缘效应(edge effect),其形成需要一定的条件,如:两个相邻群落的渗透力应大致相似;两类群落所造成的过渡带需相对稳定;各自具有一定均一面积或只有较小面积的分割;具有两个群落交错的生物类群等。边缘效应的形成,必须在具有特性的两个群落或环境之间,还需要一定的稳定时间,因此,不是所有的交错区内都能形成边缘效应。在高度遭受干扰的过渡地带和人类创造的临时性过渡地带,由于生态位简单、生物群落适宜度低及种类单一可能发生近亲繁殖,使群落的边缘效应不易形成。
发育较好的群落交错区,其生物有机体可以包括相邻两个群落的共有物种,以及群落交错区特有的物种。这种仅发生于交错区或原产于交错区的最丰富的物种,称为边缘种(edge species)。在自然界中,边缘效应是比较普遍的,农作物的边缘生物量高于中心部位的生物量。
人们利用群落交错区的边缘效应,用增加边缘长度和交错区面积的方法来提高野生动物的产量。图4-10说明了鹌鹑放养中对边缘效应原理的应用。在同样面积的土地上,种植同样量的植物,利用增加边缘长度的方法,可提高鹌鹑的饲养群数。我国南方在水网地区修造的一种桑基鱼塘,便是人类因地制宜建立的一种边缘效应,已有数百年的历史。对于自然形成的边缘效应,应很好地发掘利用。对于本不存在的边缘,也应努力去模拟塑造。随着科学技术的发展,广泛运用自然边缘效应所给予的启示,将有助于对资源的开发、保护与利用。
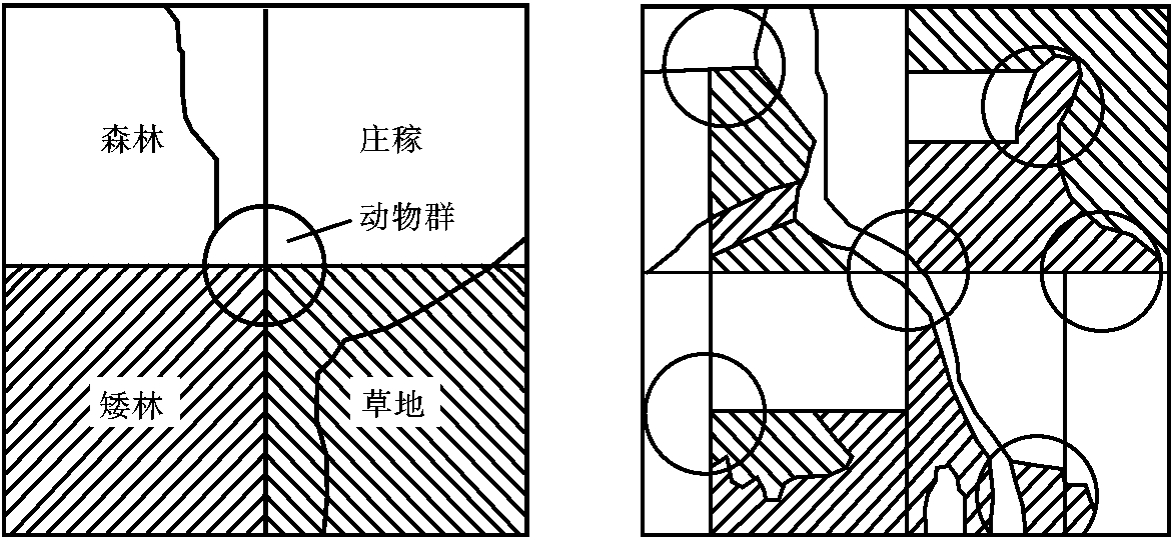
图4-10 利用边缘效应,不增加土地面积而做有效安排,以增加鹌鹑的饲养群数
(仿E.P.Odum,1959)


