第八章 微生物对环境有机污染物的降解与转化
在地球漫长的历史进程中,生物多聚物的缓慢进化和以这类物质为基质的微生物分解能力的进化是平行进行的,地球上没有任何的自然有机物过度积累就说明这一点。微生物分布广泛,代谢类型多样和适应变异能力强的特点为微生物对生物物质的巨大分解能力提供了基础。自然界物质中既有易于分解的蛋白质、脂肪、多糖,也有难以分解的木质素。环境有机污染物的进入对存在于环境中的微生物是一个挑战和选择压力。与自然存在的有机物不同,环境污染物既有自然来源,也有人工合成的,而且大多是人工合成生产的。后者也称为人造化合物(man made compounds)、人为化合物(anthropogenic chemicals)、合成化合物(synthetic compound)、合成有机化合物(synthetic organic compound)。它们中的许多其化学结构是自然界现存化合物中所没有的,因此也称为异生物源物质(xenobiotics)、异生物源有机化合物(xenobiotic organic compound)(xenote希腊文中意为外来的)。也有许多研究者把异生物源物质定义为现存酶所不认识的化合物。
环境有机污染物的导入和存在导致生物降解研究的兴起和深入。生物降解(biodegradation)被定义为生物因子(特别是微生物)作用下对物质的分解,一般来说生物降解是一个由微生物引起的衰变过程。从严格意义上说生物降解表示微生物可以完全裂解或矿化复杂的有机物成为无机物组成成分,如CO2、水和矿质成分。生物转化(biotransformation)则主要指化合物部分结构的改变。从一般意义上来说生物降解、生物转化与微生物降解(microbial degradation)、微生物转化(microbial transformation)是一致的。大部分有机污染物的生物降解发生在好氧条件下,但许多有机化合物的生物降解在厌氧条件下也能发生,但其降解速率不如在好氧条件下那样快。一些微生物也能利用化合物如NO-3、SO42-或F3+作为电子受体氧化有机物。某些化合物生物降解(如卤化烃)至少在开始时在厌氧条件时降解更快。本章把降解底物定位在有机污染物,微生物的降解作用统一使用生物降解的术语。存在于环境中的大量有机物(自然物质和部分污染物)易于被微生物所降解,可以称为易生物降解物质(readily biodegradable substances),其余的化合物(主要是污染物和少量自然物质)则难为微生物所降解,可以称为很少或抗降解物质(poorly biodegradable substances or resist biodegradable substances)和顽固性化合物(recalcitrant substances)。我们一般把这类抗降解的化合物称为难降解污染物。许多难降解污染物还具有较强的生物毒性,所以这部分污染物也被称为有毒有机物(toxic organic chemicals)或有害有机物(hazardous organic chemicals)。有机污染物的被降解程度可用完全降解(complete degradation)、部分降解(partial degradation)来表述。完全降解指有机物被氧化成CO2、水及其他无机物,并且形成新的微生物生物量。部分降解指分子的部分分解(如一个季碳原子上的脂肪链)或形成一种新的更稳定的化合物(如从简单的酚形成多酚)。与部分降解相关的是初步降解,初步降解来源于表面活性剂的降解研究,指分子的很少转化导致化合物某些特征性质的丧失,如表面活性剂疏水性脂肪(hydrophobic aliphatic)链的足够的缩短从而失去表面活性和发泡能力。部分或不完全降解的原因是;①缺乏合适的降解酶;②共代谢;③导致聚合或合成比母体化合物更稳定、更复杂的化合物;④产生毒性更强的中间化合产物。
生物降解作为生态系统物质循环过程中的重要一环,在其中起重要的作用。在C、H、O、N和S的循环中,没有微生物的活动这些元素就会被束缚在复杂的不被降解的物质中,它们就不能回到自然循环中。另外,微生物对死亡生物体的降解可以防止生物体积累在地球表面。
环境难降解污染物因其难以降解而长期残留,这就引起两个方面的问题,一是它们的分布广泛,另一个是生物富集。难降解污染物在环境中的存留时间长,因而可以输送到很远的地方。有的污染物不但难降解,而且是脂溶性的,它们在生物体中不但未被分解和大量排泄,还会贮积下来,再沿着食物链传递,使更高营养级生物比低营养级生物积累更多的污染物,从而对生物和人类造成严重的健康损害。由于每天都有大量环境污染物进入环境,填埋渗漏,空气、水、土地中有害污染物污染事件不断被披露,生物降解引起了人们的极大关注。
研究难降解污染物的降解是当前生物降解的主要课题。这种研究有重要理论和实际应用价值。一是通过研究可以查明化学结构与生物降解的相互关系,为化学家合成新的易降解有机物(环境友好材料)提供理论基础。二是可以为处理含难降解化合物的污水、废弃物以及修复污染环境提供理论指导。三是可以预测特定化合物在环境中的残留和归宿,为生态风险评价提供依据。生物降解性即一种化合物对自然生物过程的敏感性已经成为我们评价一种商业产品价值的重要标准,通过生物降解性,我们可以评价一种商品的可接受性。由于这些理由,人造化合物的微生物降解研究正成为十分活跃的研究领域。
生物降解和传统上所说的对蛋白质、多糖等有机物分解在本质上是一样的,但又有分解作用所没有的新特征(如共代谢、降解质粒等),因此可视为是分解作用的扩展和延伸。
生物降解对生物地球化学循环及维持生态系统的健康有重要的意义,主要包括:①推动元素的地球化学循环;②微生物的降解过程是生态系统中碎屑食物链的起点;③移去污染物、降低生物毒性可以维持生态系统的健康。
生物降解性和抗生物降解性是一种对立统一的相互关系,一定条件下会相互转化。降解是在一定条件下作用的结果。有机物降解作用是一个复杂的过程,难以区分酶催化生物转化反应和纯碎的物理/化学效应。这样,转化产物可能是通过多途径产生的,包括:①微生物中发生的酶催化作用;②在环境中发生于胞外的酶催化作用;③物理化学催化;④转化可以是这些原理结合的结果。例如,酶催化产生的产物可以被物理化学方式进一步转化,反过来也是一样。这个问题的进一步复杂化还在于微生物能改变环境的物理化学性质。例如,微生物活动能影响生境的pH值和氧化还原状况,这样直接导致物理化学催化的变化。灭菌这种方法常被用来区别生物和非生物过程,但事实上灭菌不仅消除了微生物的活性,也影响了调查条件下培养源的物理化学性质。灭菌的大部分技术也能导致土壤理化性质的改变。灭菌土壤失去催化转化反应能力可以或不可以指示是微生物和酶所为。虽然理化过程能导致一种化合物的转化,但矿化大多数是微生物活性的结果。最重要的,许多科学家已经成功纯化参与各种转化反应的微生物酶,因此消除了所有关于代谢生物本质的所有怀疑。由此我们可以说生物降解是所有有机污染物衰减的最重要原因。
第一节 生物降解机理、过程及影响因素
生物降解机理(基本条件)、过程及影响因素是生物降解中的基本问题,从这个问题出发有助于我们进一步理解生物降解的其他问题。
一、生物降解的机理
生物降解是微生物与降解基质的相互作用,本质上是基质的热力学和微生物生理能力相互作用的结果。这种概念可用图8-1所示。
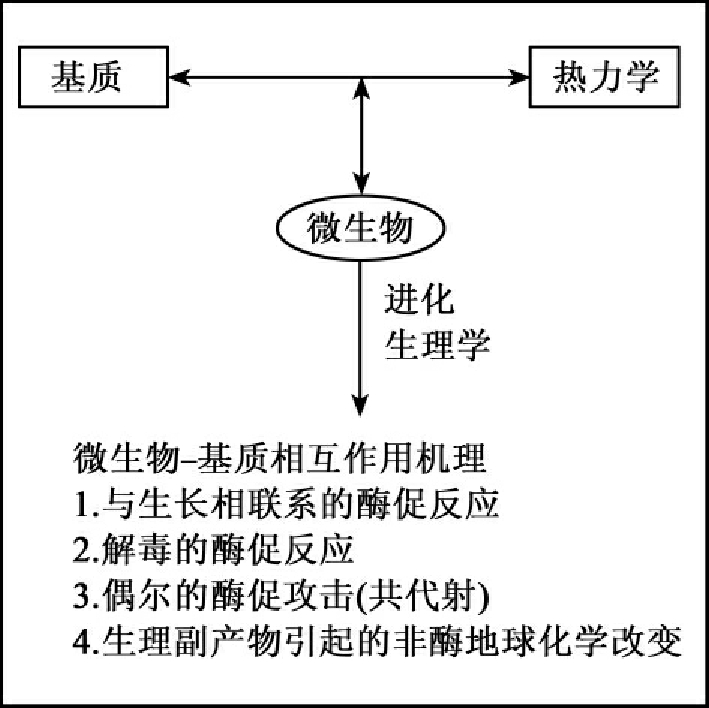
图8-1 生物降解性主要决定因子示意图
微生物生物降解过程中所催化的反应大多是一种氧化还原反应,这种氧化还原反应必须遵循一种热力学准则。图8-2图示了还原和氧化基质的相互关系,氧化-还原半反应(half reaction)的等级垂直排列。垂直轴是EH和等同的pE。半反应等级左边的化合物处于氧化态,而右边的化合物处于还原态。而且氧化态和还原态的转换受到所处系统的氧化还原态和微生物产生的酶系统催化机理的控制。高氧化条件位于图的上半部,而高还原条件排于下部。我们可以用图预示哪些半反应对的结合(combinations of half-reaction pairs)是热力学上可行的。在标准条件下下部反应产生电子,而上部反应接受电子。热力学上有利的半反应是上左下右对角线连接的反应,如光合作用的有机碳(如CH2O,图右下方)可以和存在于自然生境的最终电子受体(O2、NO-3、Mn4+、Fe3+、SO42-、CO2)连接起来。这些耦合的半反应的每一个都是由微生物所控制,碳氢化合物被氧化,而电子受体被还原。连线的长度和微生物得到的自由能成比例。微生物代谢碳氢化合物时用O2作为最终电子受体比用硝酸盐产生更多的ATP。反过来用硝酸盐作为最终电子受体又比用Mn4+和Fe3+得到更多的能量。这种模式连续下行到CO2被还原成甲烷。在反应系统中存在着半反应热力学、微生物生物学和地化学三方面的结合。值得注意的是合成卤代化合物(如四氯乙烷、多氯联苯)也存在于图8-2的等级中。这些化合物可被微生物作为最终电子受体加以利用。对卤代化合物被还原的认识有助于了解生物降解性和从环境中消除环境污染物。最终电子受体提供了可用于区分野外地点的生物化学体系和生理条件的标准。
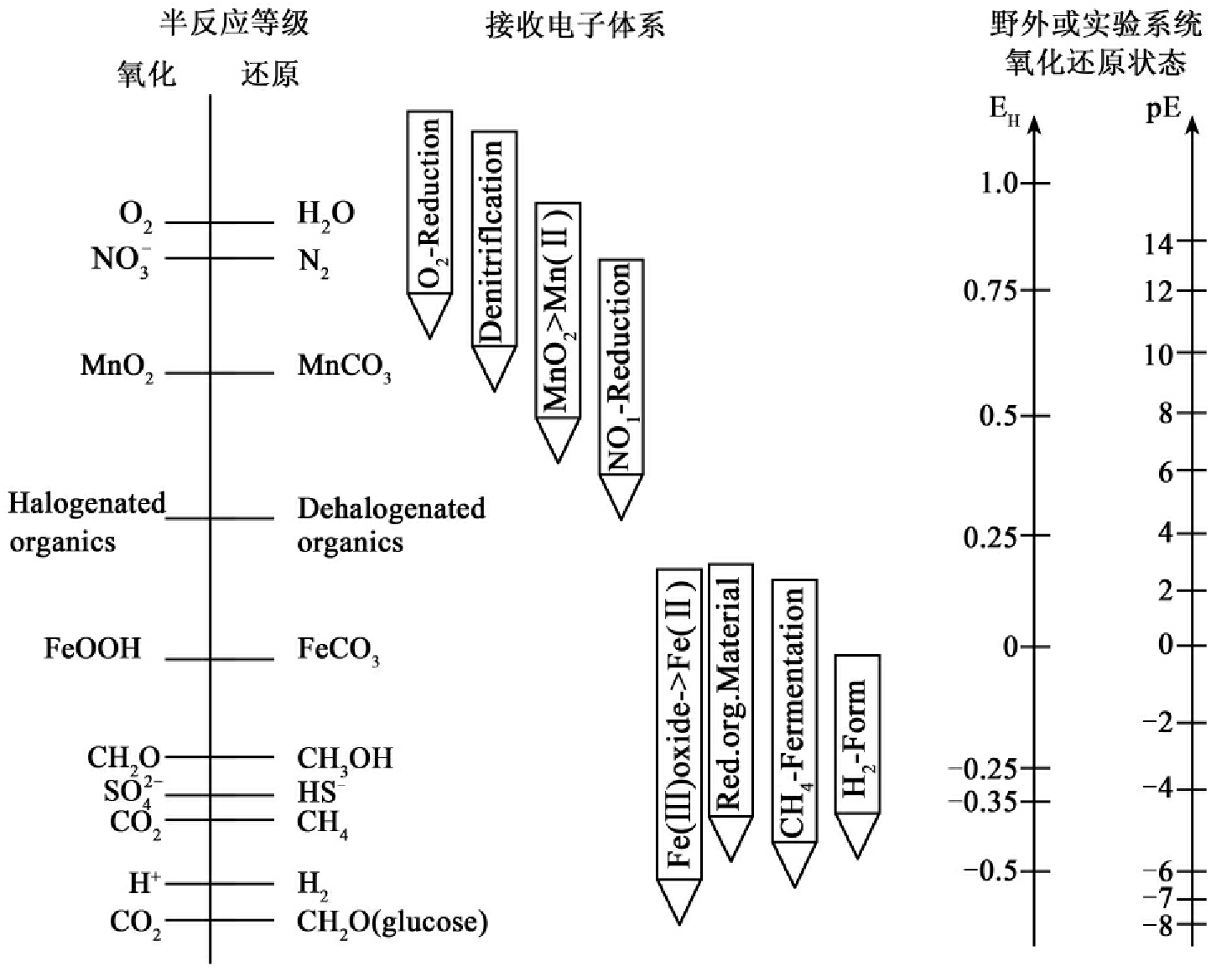
图8-2 综合生物地球化学氧化还原反应中热力学,微生物学和生理学的一种简单最终电子受体等级
二、生物降解总体过程
自然有机物的分解过程是有机物合成过程(如光合作用)的逆过程。其终产物是CO2、H2O及其他无机物。有机多聚物(如纤维素)的分解过程具有代表性,纤维素先经胞外酶降解成双糖,进入细胞再被进一步分解,微生物利用分解过程中产生的中间代谢产物及能量合成新的微生物量以生长繁殖。从这我们可以看到有机多聚物的分解过程实际上是大分子在特异性酶的作用下相关的化学键被打断,裂解出小分子物质,而后小分子物质进入细胞进一步被降解利用。环境污染物的降解过程和自然多聚物分解过程相似,也经历一个逐步降解过程。
三、生物降解的基本条件
生物降解是一个十分复杂的过程。其中细胞对底物的吸收,底物被代谢、降解和利用是最关键的环节,与这些重要环节相关的是生物可利用性、降解基因及降解酶,以及有机污染物的潜在营养性。
1.生物可利用性(bioavailability)
在生物降解过程中细胞对底物的吸收可以归结为污染物的生物可利用性。污染物的生物可利用性是其被微生物的利用能力。微生物的细胞膜是双层类脂结构。有机化合物通过细胞膜传输进入细胞内并达到的酶活性部位的过程是生物降解的基本条件。大多数微生物的活跃代谢需要高水活度(>0.96),以水溶态存在的有机物易于进入细胞,设想存在适当的代谢途径,则污染物的降解能快速进行。而低水溶性的有机物则难以进入细胞被利用。以液相存在低水溶性化合物进入水体时与水形成一个两相体系(如液态烃),微生物吸收利用这种有机物有三种模式:①溶解的有机化合物进入细胞内被降解利用;②细胞直接与有机化合物接触,或与分散在水相中的微小或亚微的液滴接触,其疏水基能溶进细胞的脂肪部分被降解,有机物以这种形式在水和化合物的界面处被逐步拉入细胞中并被代谢。微生物和不溶于水的有机物之间的有限接触面妨碍了不溶解化合物的代谢速率。加入表面活性剂(或分散剂)或微生物产生的生物表面活性剂(或分散剂)能提高低水溶性有机物的溶解度,增加其生物可利用性,从而提高吸收和生物降解的速率;③微生物细胞的疏水性表面和污染物表面的直接接触,促进吸附在细胞表面污染物的生物降解。已有研究证明选育细胞表面疏水性更强的微生物可以促进低水溶性的污染物的降解,此外产表面活性剂的菌株也可以提高细胞表面的疏水性,促进生物降解。
以液相存在的低水溶性化合物进入固相(如土壤)时还受到土壤和沉积物对其吸附作用的影响。强势(共价键)结合或进入微生物不能进入的土壤微孔后,这部分污染物将残留而不被降解。弱势(氢键、范德华力、疏水相互作用)结合仍可使污染物释放出来回到溶液中被降解。
对于固态的化合物,当其进入水相或固相介质中,微生物利用化合物的方式是直接与底物接触和吸收利用溶解态的底物。对其来说利用溶解的基质更加重要。因此低水溶性对固态有机物的生物降解的影响比液态有机物更大。
生物可利用性是生物降解的前提条件,提高生物可利用性,特别是提高难生物降解污染物(许多都是低水溶性的)的生物可利用性最关键的问题是提高其水溶性。近年来在生物修复技术中大量使用表面活性剂、乳化剂、分散剂正是要提高其溶解性,从而提高生物降解性,加速生物修复过程。
2.降解基因及降解酶
生物降解过程中的每一步都是由细胞产生的特定的酶所催化。胞外酶和胞内酶都对污染物的降解起重要作用。大分子必须在胞外被裂解成较小的亚单位以后才能进入细胞。如果没有合适的酶存在,由胞内酶或胞外酶引起的降解都会在任何一步上停止。缺乏合适的生物降解酶是导致有机污染物持久存在的一个常见原因,尤其是那些现存的降解酶不能识别的那些含有不常见化学结构的化合物。催化污染物降解的降解酶一般是特异性的,但也有一些特异性较低,如一些加氧酶,有相对较宽的底物范围,这些酶会导致污染物的共代谢。在污染物的降解酶中大部分是诱导酶,但也有组成酶,从成本效益的角度上说诱导酶要优于组成酶。
微生物的降解都是由降解基因编码的,降解基因通过转录和表达而产生降解酶,当然降解基因并不一定能表达出高酶活性,因此在生物降解中为降解基因的表达,创造良好条件也是极为重要的,如在构建降解遗传工程菌中,一般需要强有力的启动子。
3.有机污染物的潜在营养性
生物降解过程中,大部分被降解的有机污染物被用作能源、碳源、氮源、其他营养或作为最终电子受体。这种有机污染物可以认为是微生物营养基质的扩展和对易于利用基质的替代。其他一些降解(如共代谢)则不能为微生物提供能量。前者称为代谢性生产转化(metabolically productive transformation),这种降解是生产性的,可以形成新的生物量;后者称为共代谢转化(cometabolic conversions),这种降解是非生产性的,不能形成新的生物量。从微生物生理学的角度来说生产性降解是最为经济有效的。由此可以看到能给微生物生长繁殖提供能源和营养素的物质有助于微生物的生物降解。
4.生物活性
生物降解的潜力转化成生物降解的能力的基本条件是降解微生物具有生物活性,为微生物构建良好的环境条件可使降解基因得到表达,为微生物的生长和降解过程提供条件。
四、影响生物降解的环境因素
决定生物降解性的因素包括:①结构和物理状态;②化合物对生物的驯化时间;③环境条件,在决定实际的一种化合物降解时其和结构一样重要。
任何可以对微生物存活、活性及污染物状态产生影响的环境因素都对环境污染物的生物降解产生极其重要的影响。主要的环境因素包括末端电子受体、有机质含量、氮磷含量及环境温度、pH值、盐度、水活度等。
1.末端电子受体
末端电子受体主要包括氧及硝酸盐、铁离子和硫酸盐等。氧对污染物的生物降解是非常重要的,生物降解中以氧作为电子受体的氧化反应都要在好氧条件下进行,一般说来好氧的生物降解比厌氧快得多,好氧生物处理比厌氧生物处理有高得多的效率。例如进入淡水湖泊和河流好氧区的石油类烃一般对微生物降解是敏感的,但积累在厌氧沉积物中的石油一般是相当持久的。在缺氧条件下,硝酸盐、铁离子和硫酸盐可以代替O2作为可利用的末端电子受体,氧化降解苯、甲苯、苯甲酸盐等一类芳香烃化合物。
2.有机质含量
环境污染物的生物降解需要一定量的微生物,而微生物的数量要靠有机物含量来支持。高有机质含量可以支持一个庞大的微生物群落,这种微生物群落对外来污染物具有很强的降解能力。对于只能以共代谢方式进行降解的某些污染物,只有提供充足易于利用的初级基质,才能产生足够的降解污染物的酶。这种情况下提供充足的有机质特别重要。
3.氮磷营养
氮磷营养对环境污染物的降解有重要影响。微生物在利用有机物特别是利用主要由碳和氢组成的烃时也消耗像氮和磷这样的主要营养物。有研究者在监测一个温带湖泊中烃降解的季节变化时,发现这种变化受氮和磷可利用形式的控制。降解的最大速率出现在可利用氮磷含量高的早春,但在这些营养物被快速消耗后降解速率降低,而投入氮磷后又使降解速率提高。在一个降解系统中碳、氮、磷的平衡供应是十分重要的,微生物生物量平均碳氮比约为5∶1~10∶1,一般降解系统的碳(BOD)氮磷之比约为100∶10∶1。然而在某些情况下可以使用相当不同的比率。有研究发现碳氮比达到200∶1,碳磷比达到1000∶1仍可有效降解(修复)土壤中的烃。为什么要使C∶N和C∶P比值高于细胞的比值?这是因为有机碳的代谢过程中大量的碳被转化为二氧化碳释放掉,使得系统内的碳大量损失,与此相反氮、磷都掺入微生物量而保留在系统内。
4.温度
温度对微生物的生长以及环境污染物的溶解度可以产生很大的影响。微生物及生物降解可以在很宽的温度范围内进行,但中温、高温条件下生物降解速率要比低温高得多。
5.pH值
环境介质的pH值对微生物生长、代谢活性以及环境污染物的溶解性都产生极大的影响。尽管极端的嗜酸、嗜碱微生物可以在极端pH值条件保持很高的活性,但总体上说中性pH值或稍偏酸、偏碱条件下,有机污染物的降解速率最快。
6.盐度
嗜盐微生物可以在高盐度条件下生长,但非嗜盐微生物的生理活性却很容易受到盐度的影响。研究表明向淡水沉积物样品加盐后烃降解速率降低。
7.水活度
任何微生物的最适生长都需要适合水活度,水活度对环境污染物的降解也产生很大影响。高水活度有助于大多数微生物的生长,也有利于环境污染物的溶解。研究说明在高水活度条件下污染物的生物降解具有较高的速率。
第二节 微生物降解有机污染物的巨大潜力
微生物作为分解者具有对自然有机物的巨大分解能力,自然界没有大量的有机物积累就充分说明这一论点。大量的有机污染物(其结构与自然界化合物不同)进入环境是对微生物适应进化能力的一种机遇与挑战,在新的选择压力下,微生物群落也能或慢或快地进化出能降解这些化合物的酶系,获得对它们的降解能力。这样微生物就具有对所有有机物的降解能力。Alexander形象地把这种一般性的认识概括为“微生物的绝对可靠性原理”(priniciple ofmicrobial infallibility)。此外微生物在个体特征、生态分布、生理功能、遗传变异方面的诸多优势,使微生物具有对环境污染物降解的巨大潜力。
一、微生物分解自然有机物的能力良好基础
存在于地球表面种类繁多、数量庞大的无机(如SO2、NH3、H2和CH4)和有机(碳水化合物、脂肪、蛋白质、核酸和烃)物质被扩散到同样多样的生境。生境的物理化学特征异质性强,包括pH值、温度、盐度、氧压、氧化还原电位、水势等。这些多样的化合物和多样环境是微生物进化过程中的选择压力源。微生物进化的结果是利用这些化合物生长和存活。微生物的生长和存活推动了元素的生物地球化学循环,并为地球上其他生物的栖居提供了条件,深刻了解和认识微生物在维持生态系统中的作用提供了从理论上和实践上研究微生物对有机污染物降解过程的重要思路。
微生物应对自然有机物的存在而进化出生物降解活性。这些有机物包括多聚物、腐殖质和如CH4的C1化合物。了解这些天然有机化合物的归宿是重要的,这是因为针对这些化合物进化出来的降解活性是降解处理的基础,这种处理可用于解决于泄漏到环境中的有机污染物的问题。
有机多聚物包括植物多聚物、构成真菌和细菌的细胞壁多聚物及节肢动物骨骼多聚物,三种最常见的多聚物是植物多聚物纤维素、半纤维素和木质素。还有其他多种多聚物,包括淀粉、几丁质、肽聚糖。按结构分两类,以糖为基础的多聚物,包括环境中大多数多聚物,及以烷基苯为基础的多聚物(如木质素)。纤维素是最丰富的植物多聚物,也是地球上最丰富的多聚物,它是由β-1,4葡萄糖亚单位连接而成的线性分子,每个分子含1000~10000亚单位,分子量高达8×106U。纤维素分子量大,不溶于水。微生物对纤维素的降解是由胞外酶和胞内酶结合进行。胞外酶是β-1,4葡聚糖内切酶和β-1,4葡聚糖外切酶。内切酶在多聚物内随机水解纤维素分子,产生越来越小的纤维素分子。外切酶连续地从纤维素分子还原性末端水解出两个葡萄糖亚单位,释放出纤维二糖。纤维二糖被纤维二糖酶(既是胞外酶也是胞内酶)水解成葡萄糖,纤维二糖和葡萄糖都可被细菌和真菌吸收。
其他植物多聚物半纤维素、淀粉、几丁质的降解和纤维素相似,但酶类组成及过程会有一些不同。
木质素在结构上和所有以碳水化合物为基础的多聚物不同。木质素的基本结构是两种芳香族氨基酸——酪氨酸和苯丙氨酸。它们被转化成苯丙烯亚单位如香豆醇、桦柏醇和芥子醇。500~600个苯丙烯亚单位随机聚合,结果形成无定形的芳香族多聚物木质素。同其他有机多聚物相比,木质素的生物降解较缓慢且不彻底。木质素降解缓慢是因为是其由高度异质的多聚物构成,且含比碳水化合物残基更难降解的芳香族残基。分子的巨大异质性,妨碍进化出可与纤维素降解相比的特异性降解酶。作为替代,一种非特异性胞外酶依赖H2O2木质素过氧化物酶与一种能产生H2O2的胞外氧化酶结合。过氧化物酶和H2O2系统产生氧自由基,自由基与木质素多聚物反应释放出苯丙烯残基。这些残基被微生物细胞吸收和降解。
苯丙烯残基是自然界中的芳香族化合物,其结构同几种有机污染物分子如BTEX(苯、甲苯、乙苯、二甲苯)、多环芳烃、杂酚油相似。实际上苯丙烯的降解途径与芳香族化合物降解途径非常相似。深入研究过的能降解木质素的自腐真菌原毛平革菌(Phanerochaete chrysosporium)也能降解结构上与木质素相似的污染物。
二、微生物降解有机污染物的潜力
有机污染物是一个很难界定,而内涵又十分复杂的概念。对一种具体的化合物而言,在不同的场合,不同情况下会得到不同的结论。但从化合物的结构上说部分有机污染物结构和自然界有机物结构相似,部分结构与自然基质结构部分相似,有的其结构与自然基质结构差异极大。
从总体上说微生物对自然有机物和有机污染物的降解模式是一致的,本质上降解途径的每一步都是由细胞产生的特定酶所催化。大分子必须在胞外被裂解成较小的亚单位才能进入细胞。生物降解过程的前提是现存酶能否认识这种化学结构。这样在环境条件能满足微生物生长的正常情况下,那些结构与自然基质结构相似的有机污染物易于降解,如石油烃中的链烷烃。对结构上与有机基质不同的有机污染物,微生物一时不能降解它们,但微生物群落能通过基因突变、接合作用、转化及转座造成的基因转移和重组而获得生物降解能力。已经从自然环境中分离到大量能降解大量结构上与常见有机物完全不同有机物的微生物,DDT、2,4,5-T这样难降解污染物的降解菌都已从土壤中分离出来。这就是说微生物可以通过学习和进化而获得对各种异生物源化合物的降解能力。从某种意义来说,生物降解是绝对的,而不降解却是相对的。
此外即使微生物无法进化出任何的降解能力,但微生物中存在一种共代谢能力,许多微生物的降解酶具有广基质专一性,使许多降解酶在降解其专一性的底物的同时能共代谢结构相似的污染物,从而从水平方向上扩大了降解范围,促进生物降解。一种微生物的共代谢降解不会使一种污染物被完全降解,但微生物群落众多不同种类的微生物却可以使污染物被完全降解。
和有机污染物的广泛扩散和分布相匹配的是降解生物(主要是降解微生物)的无处不在。现在已有大量的可以降解各类污染物的微生物从土壤、水体及其他环境介质中分离出来。主要是细菌和真菌。细菌中最重要的是好氧革兰氏阴性细菌,包括假单胞菌属、鞘氨醇单胞菌属、伯克霍尔德氏菌属、产碱菌属、不动杆菌属、黄杆菌属、甲烷氧化菌、硝化细菌。革兰氏阳性细菌包括节杆菌属、诺卡氏菌属、红球菌属和芽孢杆菌属。某些反硝化细菌、硫酸盐还原菌和甲烷产生菌也参与有机物的厌氧降解。参与污染物降解的真菌主要有平革菌属(白腐真菌)、青霉菌属、曲霉属、木霉属和镰孢。平革菌属的某些种能够降解结构上极为复杂的污染物,如农药DDT、多环芳烃、木质素等。
三、微生物降解有机污染物的优越条件
微生物在降解有机污染物方面在形态结构、生长繁殖、生态分布、生理功能、遗传潜能群落组成、迁移能力诸方面具有巨大的优势,这些保证了微生物对有机污染物降解的巨大潜力。
1.体积小,比表面积大
微生物在形态结构上具有个体微小(small size)、高比表面积(high specific surface area)的特点。这一特点使微生物能与环境介质及其中的环境污染物密切接触,使微生物能进入所有存在环境污染物的位置,并有助于环境污染物扩散到细胞内,有助于微生物对环境污染物的降解。
2.生长繁殖快
微生物在生长繁殖上具有潜在的高生长速率(potentially rapid growth rate)的特点。微生物在不同条件下的繁殖速度差异极大,高速生长时18分钟可以分裂一次,而慢时繁殖一次则需要数月以至更长。这样在大量有机污染物存在时,微生物能以较快的生长速率生长,并可达到较高的生物量浓度,从而为处理高浓度污水,废弃物,修复污染环境创造条件。
3.分布广泛
微生物在生态环境中无处不在的生态分布(ubiquitous distribution)是微生物净化环境的最大优势。微生物在自然的生态环境中分布极为广泛,江河湖海、土壤矿层、大气上层以及人体、动植物几乎无处不有微生物的存在。甚至在高等生物及其他生物不能生存的极端环境下也有微生物的存在。微生物的这一特点为利用微生物消除污染物,净化污染环境提供了良好条件。很多难降解的环境污染物由于长时间存留在环境中,因而可以随大气、水流而传播到离使用地点很远的地方。有研究表明南极冰雪中存在DDT,那里也同样存在相当数量的微生物,那里的DDT因而也被降解。
4.代谢多样性
微生物在生理功能上具有代谢多样性和高代谢活性潜力(potentially high rate ofmetabolic activity)特点,这些特点使微生物在环境污染物面前表现出极高的生理活性。微生物代谢多样性表现为多样降解酶、多样降解途径和对污染物的多样利用方式。微生物在适应自然有机物的过程中进化出非常多样的降解酶,既有胞内酶又有胞外酶,既有降解利用小分子化合物的酶,也有分解大分子的酶,微生物的所有的酶都不同程度地介入环境污染物的降解过程。六大酶类都可以发挥作用,氧化还原酶类、转移酶类、水解酶类、裂合酶类、异构酶类、合成酶类都出现在降解过程中。有的是结构酶,有的是组成酶。微生物在降解有机物的过程中还表现出多样的特点,不同的微生物种类,甚至不同的菌株对同一有机物可以有不同的代谢途径,甚至在不同条件下同一菌株也可有不同的代谢途径。微生物对环境污染物还表现出不同的利用方式,有的作为唯一碳源及能源;有的是共代谢,不从降解过程中得到能源。此外微生物在营养丰富,环境条件适宜的条件下具有极高的代谢活性,其利用和合成有机物的以体积计的代谢活性要比其他动植物高出千万倍。微生物灵活多样的生理特征使微生物面对多样的污染物而能做出不同的生理反应。
5.易变异,适应能力强
微生物具有遗传方面的变异,适应能力强的特点,为微生物进化出对新的环境污染物的降解能力提供了良好条件。没有任何生物的变异能力比得上微生物,可以很容易得到微生物抗性菌株、缺陷型菌株,而且微生物的遗传背景是研究得最清楚的。微生物的自发突变、诱发突变易于发生,遗传重组的途径多样,这为微生物从遗传信息角度适应异质的污染环境和污染物的选择提供了基础,此外微生物还有丰富的降解遗传信息,除染色体(细菌为似核)编码降解酶外,核外遗传物质——质粒也可以在降解中起重要作用。
6.种类多,数量大
微生物在群落结构上具有种类繁多,数量庞大的特点,这种特点为微生物降解各种各样的污染物提供了物质基础。种类繁多,数量庞大的微生物群落使其能经受得起各种环境压力,即使在几乎灭绝的条件下仍有少数能生存下来,适应新的环境而能生长繁殖下来能对污染物进行降解,另一方面庞大群体可以保证即使在极低的突变率时也有一定数量个体发生突变,并且由于环境的选择而使这种有利于降解的突变保存并逐步发展起来。
7.微生物的迁移能力
微生物在生态环境中的迁移可以通过主动运动和被动迁移而获得。这种迁移可以使降解菌从一个地方迁移到另一个地方,使它们到达被污染环境。在生物修复中使外源降解菌迁移到作用位点是提高生物修复能力的重要问题。
四、生物降解作用的认为强化
大量研究工作表明微生物降解环境污染物的潜力远未充分开发利用,对现有降解菌的生理遗传改造,还可以进一步提高这种降解能力。生物降解作用的强化提高是生物降解中的重要研究课题。提高生物降解能力的方法从组织层次上包括:①群体降解水平的提高,如向环境投入营养物可以从总体上提高微生物的生理活性。②微生物种群降解能力的提高,通过对降解微生物的生理遗传改造,提高降解能力。这包括生理层面的驯化适应,遗传层面的遗传修饰改造及遗传工程改造。③酶工程的降解酶改造扩大酶底物范围,提高降解能力。
从种群水平的工作程序可以包括对现存降解能力的提高以及构建新的降解能力。
1.现存生物降解能力的提高
(1)自然生物降解多样性的分析和解降能力资源开发利用
自然发生的微生物活性过去是,现在也是所有生物技术应用的出发点和基础。分离具有新的代谢能力的细菌菌株,并对它们的降解途径作出生物化学和遗传学阐述,这样有利于开发自然发生的多样性和降解能力资源。主要的应用方面是可以克隆遗传基因构建新的遗传工程菌以及构建协同式菌群,构建畅通的代谢降解路线。
在芳香化合物生物降解中,各种双加氧酶的羟化作用可以增加化合物的极性,促进生物降解,因此在这类化合物降解启动有重要作用。导入的两个羟(基)氢氧基(hydroxylgroup)位于邻位或对位,完成催化过程的酶是多成分的双加氧酶,酶是由一个电子传递链和具有催化活性的α-和β-亚基组成的。这类酶的多样性和底物范围受到特别关注。鞘氨醇单胞菌(Sphingomonas sp.)RWI菌株产生的二(dioxin)双加氧酶是最先被报告的酶,其能进行一种成角(度)(angular)的使一对邻近的碳被氧化的双氧化,其中的一个碳被包含在两个芳香环之间桥的之中(one of which is involved in one of the bridge between the two aromatic rings)。编码二双加氧酶的基因也有独特的特点,一般的双加氧酶的编码基因成簇,而这种酶却非预期地分散于染色体。另一个能进行成角(度)双氧化的咔唑(carbazole)1,9α-双加氧酶(in Pseudomonas sp.strain CA10)也有其特点,其末端的加氧酶由单一蛋白质CarAα组成,和典型(classical)的由大的α和β亚基组成的加氧酶不同。CarAα的核苷和推想的氨基酸序列也是独特的,与其他的末端加氧酶(包括二双加氧酶)的大亚基仅展现出很少的相似性。对羟化作用的大量双加氧酶的研究说明环激活的双加氧酶存在着广泛的多样性,从这个角度出发,我们还可以发现具有新的特异性的基因。
细菌降解多样性不限于环激活的双加氧酶,也包括降解芳香烃化合物代谢产物的酶。芳香环断裂是卤代芳香烃降解的主要关键反应,其降解酶的多样性也广受关注。长期以来认为通过间位(meta)裂解途径代谢3-氯代儿茶酚是不可能的,这是由于反应产物将失活二醇外(extradio)双加氧酶。但恶臭假单胞菌GJ31菌株却含有一种新的氯代儿茶酚2,3-双加氧酶,这种酶能有效地打断3-氯代儿苯酚(在2,3-位置),这导致同时环断裂和脱氯,细菌能通过间位断裂途径降解氯代苯的途径已被详细研究,抗自杀失活的残基已被定位。五氯酚和γ-六氯环己胺(林丹)也以同样方式被降解。
参与各种硝化芳烃化合物降解的新的类型的间位断裂双加氧酶也有报告。环裂解的基质通常是二酚,二酚中的二个羟基不是邻位(ortho)就是对位(para)。在代表性反应中,仅一个羟基存在于环断裂基质2-氨基酚中,其经历环断裂到2-aminomuconic半醛,氨基明显替代第二个羟基功能。
(2)表达生物表面活性剂提高异生物源化合物的生物可利用性
环境中的疏水性有机化合物长时间抗生物降解的主要原因是它们的生物可利用性受到溶解性的限制。提高它们的生物可利用性,从而促进生物降解最可能的方法是应用(生物)表面活性剂。大量的研究都表明导入表面活性剂可以促进生物降解。然而表面活性剂在生物修复中的有效性的报告是混淆不清的(mixed)。生物表面活性剂的本质作用已被解释为可以增加疏水性、水不溶性生长基质的表面积,通过增加溶解度或从表面上解吸下来,以及调控微生物对表面的吸附及脱离从而增加化合物的生物可利用性。这样加入表面活性剂对生物降解的净效率将是化合物的溶解度增加和细菌对这些化合物直接吸附降低的综合结果。表面活性剂的加入会降低细菌吸附到非水液体表面,这样降低了在疏水性化合物上的生长。实际上表面活性剂的相应效应我们还了解很少,是极其复杂的不同环境中微生物和土壤、沉积物、污染物相互作用的结果,所以在使用单一表面活性剂时往往有不同的效果。这样我们对这些相互作用还有待进一步研究了解,在使用表面活性剂时要优化表面活性剂,了解生物降解和靶环境条件的相互作用。产生生物表面活性剂的具有降解能力的菌株将有助于生物降解,将是一种最好的选择。
(3)提高细胞吸收有机污染物能力,促进生物降解
许多有机污染物要进入细胞内才能被胞内酶降解,因此把有机物运输进入细胞对生物降解也是十分重要的。许多芳香化合物可以通过能量依赖迁移系统被细菌吸收。Pao等指出有三类(three families)透性酶负责酸性芳烃化合物及其代谢产物的转运。新的研究资料表明异生物源化合物也是被特异的运输系统转运,Leveau等首先报告了一种转运2,4-二氯苯氧醋酸盐(2,4-dichloro phenoxyacetate)的运输系统(transporter)。对手性化合物(chiral)的对应选择(enantioselective)吸收也已得到证实,除草剂2-(2,4-二氯苯氧基)丙酸盐就是这样的结构,其被Sphingomonas herbicidovoransMH降解时就存在三种可诱导的质子-梯度-驱动吸收系统(inducible,proton-gradient-driven uptake system)。在降解蒽的荧光假单胞菌中也存在对非电性的(noncharged)疏水性蒽分子的主动运输系统。这些研究结果说明我们为修复而设计超级生物降解菌(或称为超级生物催化剂superior biocatalysts)时也必须考虑对污染物的吸收机制。
(4)增强对有机污染物的趋化性,促进生物降解
微生物对污染物的趋化性也可以增强发生在自然环境中的生物降解。已有研究证明恶臭假单胞菌的4-羟基苯酸盐的运输系统pcak也使细菌对这种化合物的趋化性起作用。其他许多运输系统(如透性酶)也能起到决定趋化性的化学受体作用,它们和已描述的化学受体是不同的。Grimm和Harwood报告说,恶臭假单胞菌的降解质粒(NAH7)上的nahy基因编码的膜蛋白Nahy也决定着对萘的趋化性。它的羟基-末端区类似于趋化性的转换器(transducer)蛋白,因此具有对萘化学受体的功能,其也可以成为像联苯这样相关的化合物的化学受体。对趋化性遗传基础的研究将使我们能够利用趋化性的特征来增强生物降解作用。
(5)改变细胞表面特征提高适应环境和生物降解能力
许多高度疏水性的环境污染物(如甲苯等)对微生物具有很强的毒性,其可以积累在细胞膜上,干扰细胞膜,使细胞失活,从而就会阻碍所希望的生物降解。具有降解能力的微生物也会因细胞膜受损而失去其降解功能。现在已经发现对有毒污染物的耐受能力是它们具有降解能力的重要因素。
耐受能力的原理主要有三种。
①细胞膜组成成分中脂肪酸从顺式(cis)转变成反式(trans)的异构化可以提高耐受能力。反式脂肪酸较高的刚性使膜结构对有机溶剂的干扰敏感性较小。编码使脂肪酸从顺式异构化成反式的酶的基因已被克隆。使溶剂耐受性增强的微生物磷脂生物合成量增加的现象已被观察到。
②通过修饰外膜蛋白和脂多糖来降低细胞表面的疏水性,从而减少有机溶剂在细胞膜上的积累,这从另一个角度提高耐受性。
③微生物存在溶剂泵出系统,从而提高耐受性。
许多分离出来的溶剂耐受细菌已被证明能降解矿化有机溶剂,它们的代谢能力已被用于工程化的生物修复。在许多芳烃严重污染的地方,溶剂耐受菌成为最先的定殖菌,成为这些污染物去除的优势菌。使用有降解能力的溶剂耐受菌可以成为生物修复有前途的方法。
对有机溶剂的这种耐受性我们实际上可以理解为一种适应,一种对环境的适应。适应环境是生物降解的前提条件,有人已构建出重组子耐放射异常球菌(Deinococcus radiodurans),其能在高辐射环境下氧化甲苯、氯苯等污染物;用于极端环境修复的极端降解菌已得到应用,例如在北极和南极地区,降解PCB的耐冷细菌已分离出来。细胞表面的疏水性的改变不仅代表一种对有机溶剂的防御机制,同时也对细菌表面吸附产生影响。在生物修复中,我们希望外源接种细胞不是堵塞在周围而是迁移到所希望的地方。黏附缺陷型菌株在运动迁移上具有优势,现在能在土壤中快速迁移的突变株已被筛选,并已得到实际应用。
2.发展新的生物降解能力
(1)构建降解型基因工程菌
构建组合式、互补式降解能力的新菌株可以大大提高微生物的降解能力。构建超级生物催化能力的一种策略是把源于不同生物的降解片段合理组合并转移到一种受体菌株,从而在对异生物原物质生物降解中避免形成截止式产物或毒性更强的代谢产物,因而达到完全的降解过程。这种策略已被成功用于降解高毒性化合物三卤代丙烷(trihalopropanes)。例如Bosma T等先构建一个广宿主范围质粒(broad host-range plasmids),这种质粒含有编码卤代(键)烷烃脱卤酶的基因(来自Rhodococcus sp.Strain M15-3),可在不同的异源启动子控制下实现表达,这种酶能有效地把三卤丙烷转化成二卤丙醇。
有研究者在氯苯酸盐降解途径上结合一段氧化途径,这种途径能把(氯)二苯转化(由bph基因编码酶进行)成(氯)苯酸盐。实现这种组合的方法是通过细胞的接合融合得杂合菌株,或把bph基因转入氯苯酸盐降解菌。Hrywna Y等克隆出氯苯酸盐邻位和对位脱氯酶的编码基因,并在降解二苯和共代谢氯代二苯菌株(睾丸酮丛毛单胞菌Comamonas testosteroni strain VP44)中表达。结果表明构建的工程菌株能生长并对2-和4-氯二苯脱氯降解(构建基因工程菌见本书第十五章)。
(2)构建转基因植物,提高植物的净化能力
植物已被广泛用于污染环境的生物修复。把来源于细菌的遗传信息转入植物将更有利于提高植物的净化能力。转基因植物白杨(poplar plantlets)能表达细菌的汞还原酶,已经证明能在离子汞表现出毒性的水平下萌发和生长,并能释放出元素汞,因此能有效地把土壤中的结合汞排出。同样转基因植物能表达修饰的有机汞裂解酶,它们能旺盛生长在较高浓度的高毒性有机汞条件下,有机汞裂解形成的离子汞会积累在可任意利用的植物组织中。阴沟肠杆菌PB2(Enterobacter cloacae PB2)能以TNT作为N源生长,其表达的季戊四醇四硝酸盐(pentaerythritol tetranitrate)还原酶能还原TNT的芳香环,并释放出亚硝酸盐。把细菌这种酶的基因转入烟草,其种子也能表达这种还原酶,也能在丙三氧(基)三硝酸盐或TNT毒性浓度条件下萌发和生长,而此时野生型种不能萌发和生长,其幼苗也比野生型幼苗对丙三氧(基)三硝酸盐有更快、更完全的脱硝能力。实际上转基因植物在生物修复中可以作为细菌的替代物。
(3)降解酶的定向改造和进化,提高降解能力
酶是生物化学反应过程的核心,各种降解酶也是生物降解过程中的关键因素。研究酶基因的克隆和表达、酶蛋白的结构和功能的关系以及对酶进行再设计和定向加工的基因工程、蛋白质工程方法和技术的发展及进步为发展更优良的新酶或新功能酶提供了广阔的前景,这也为降解酶的发展提供了新的技术手段。
酶分子本身蕴藏着很大的进化潜力,许多功能有待开发。目前酶工程主要采用基因定点突变(site directed mutagenesis)和体外分子定向进化(in vitro molecular directed evolution)两种方式对天然酶分子进行改造。
1)基因定点突变。基因定点突变的基因突变是在了解酶蛋白的三维空间结构及编码序列,搞清结构与功能关系的基础上,根据蛋白质的空间结构知识来设计突变位点,然后通过点突变使已知的DNA序列中一定长度的核苷酸片段发生替换、插入或缺失。而改变蛋白质结构中特定位置(如活性中心)的氨基酸残基,从而改变酶的特性,最后通过筛选选出有益的突变从而提高酶的活性或开发了新的酶。目前已利用定点突变技术改进天然酶蛋白的催化活性、抗氧化性、底物特异性、热稳定性及拓宽酶反应的底物范围,改进酶的别构效应(allosteric effect)。这种方法与使用化学因素,自然因素导致突变的方法相比,具有突变效率高,简单易行,重复性好的特点。
点突变的方法及技术也可以用于降解酶的改进,并且已显示重要的应用前景。卤代烷烃(haloalkane)脱卤酶是异生物源化学物降解中最先受到关注的酶,为了详细了解不同卤烷烃脱卤酶的专一性,蛋白质序列和酶的三维结构模式已被进行了比较,功能上重要的氨基酸被指出,这些氨基酸可以作为将来定位诱变实验的目标。根据已知的三维结构,Vollmer等构建了一个黏康酸盐异构酶(muconate cycloisomerase)(参与自然芳香化合物的降解)的变异株,其结合穴(binding cavity)中的氨基酸也见于氯(代)黏康酸盐异构酶(chloromuconate cycloisomerases)(参与氯化芳香烃的降解)的相同位置,并能增加某些氯代黏康酸盐的特异性常数(specificity constants),然而在许多其他方面,突变体酶保留了野生型的特点,这说明不论结合穴是简单改变,还是更复杂的改变都可以造成酶的可见差异。
在不能取得结构信息时,对酶之间的氨基酸序列所作的对比分析能找出对催化活性有重要作用的氨基酸残基。通过具有不同底物专一性或催化特点的相关酶的杂合体(hybrids)的分析,可以得到关于酶决定基质专一性的酶区域(regions)的信息。这种方法已被用于查明联苯双加氧酶底物专一性差异的相应的残基。Parales等的研究表明2-硝基甲苯2,3-二加氧酶的α-大亚基的C-末端区域决定的酶的专一性。BeilS等研究甲苯和氯苯双加氧酶产生的杂合酶(hybrid enxyme),结果证明甲苯双加氧酶活性中心(active site)附近一个氨基酸从Met220(甲硫氨酸)转换成Ala(丙氨酸)能使酶对1,2,4,5-四氯苯转化和脱氯。
有研究结果说明二苯双加氧酶(伯克霍尔德氏菌CB400菌株)的底物专一性是由活性中心的三个区域决定的。ZielinskiM等进一步研究这些区域中的23个氨基酸的效应。结果证明替代这些氨基酸会直接影响与底物的相互作用,另一方面也证明不与底物相接触的许多氨基酸(Ile 243,Ile 326,Phe 332,Pro 334,and Trp 392)也强烈改变中心双加氧作用的表现。这说明预测中没有影响的氨基酸残基也可以对专一性起关键作用。这样应用定位突变可以使一种单一的酶扩大其生物降解基质的范围。
2)体外分子定向进化。酶的体外分子定向进化是在人工模拟自然进化过程的条件下,通过容错PCR、DNA改组、交错延伸、随机引物引导重组和递增截短等方法对编码酶的基因进行随机突变和体外重组,经高通量筛选获得性能更优良或全新的酶。
酶定向进化通常分三步进行:第一,通过随机突变和(或)基因体外重组创造基因多样性。第二,导入适当载体后构建突变文库。第三,通过灵敏的筛选方法,选择阳性突变子。这个过程可重复循环,直至得到预期性状的酶。其中获取多样性基因是整个工作的基础,是酶的定向进化成功的关键。
①基因随机突变。容错PCR(error-prone PCR)技术、化学诱变剂介导的突变、致突变剂产生随机突变和随机寡核苷酸突变可以造成基因随机突变。容错PCR技术是一种相对简单、快速廉价的随机突变方法,通过改变PCR反应条件,使扩增的基因出现少量碱基错配,从而导致目的基因的随机突变。化学诱变剂(如羟胺)直接处理带有目的基因片段的质粒也可产生随机突变,然后用限制性内切酶切下突变的基因片段,克隆到一定的表达载体中进行功能筛选。致突变株体内的DNA突变率比野生型高出数千倍,将带有拟突变基因的质粒转化到致突变株内培养,也可以产生随机突变,频率一般为1/2000。
②基因体外重组。体外重组有同源基因重组(homology-dependent gene recombination)和非同源基因重组(homology-independent gene recombination)。同源基因重组在体外重组中占有重要地位。
a.同源基因重组。同源基因重组的主要方法包括DNA改组、家族DNA改组(family DNA shuffling)、交错延伸过程(staggered extension process,StEP)、随机引导重组(random priming recombination,RPR)等方法。DNA改组系将一群密切相关的序列,如多种同源而有差异的基因(或一组突变的基因文库),在DNaseⅠ的作用下随机切成小片段,这些小片段可通过自身引导PCR(self priming PCR)延伸并重新组装成全长的基因。这些重排产物的集合又称为嵌合文库(突变文库)。再对嵌合文库进行筛选,选择改良的突变体。家族DNA改组(也被称为自然发生的同源序列的shuffling或DNA shuffling)采用一系列天然存在的同源性较高的基因作为起始基因进行DNA改组操作。此法存在重组子产率低的问题,解决这一问题是减少亲本背景,人们进行了两项改进,以单链DNA(ssDNA)替代双链DNA(ds-DNA)或用限制性内切酶取代DNaseⅠ。由此形成的DNA片段没有交错重复,从而减少了同源双链(home duplex)的形成,提高重组频率。这种方法已在降解酶定向进化中得到应用。交错延伸过程是在DNA改组的基础上发展起来的简化的DNA改组方法。这种方法将含有不同点突变的模板混合,短暂地进行退火(annealing)及延伸反应;在每一轮中,那些部分延伸的片段根据序列的互补性与不同模板退火并进一步延伸,反复进行,直到获得全长基因片段。随机引导重组利用随机序列引物产生大量互补于不同部分模板序列的DNA短片段,由于碱基的错误掺入或错误引导,这些DNA片段中也含有少量的点突变,在随后的PCR反应中,DNA小片段可相互同源引导和扩增成全长基因,然后克隆到适当的载体上表达并通过适当的筛选系统加以选择。
b.非同源基因重组。非同源基因重组不要求酶基因序列的同源性。主要的方法是递增截短法(incremental truncation),这种方法的核心是以核酸外切酶Ⅲ代替DNaseⅠ对靶序列进行切割,由于其5'-3'外切核酸酶的作用,因此得到的递增截短片段库(incremental truncation libraries,ITLs),理论上包括了靶序列DNA单碱基对删除的各种情况,使得在较低的复性温度下,可实现非同源序列间发生重组。依靠这种方法可获得不依赖DNA序列间同源性的杂合酶。
c.选择和筛选。当突变体酶赋予细胞生长或存活优势,赋予寄主对药物的抗性或可满足营养缺陷型菌株的生长需求时,可容易地从有106以上的酶蛋白突变体的文库中筛选出所需要的酶。
d.家族DNA改组在降解酶定向进化中的应用。联苯双加氧酶(BphDOX)催化联苯和相关化合物开始的氧化。这种酶是一种多组分酶(multicomponent enxyme),其中的一个大亚基(由bphA1基因编码)决定基质的专一性。Suenaqa A等以类产碱假单胞菌(Pseudomaonas pseudoalcaligenes KF707)KF707菌株和洋葱伯克霍尔德氏菌LB400菌株(Barkholderia cepacia LB400)的bphA1作为同源基因进行DNA shuffling,得到许多定向进化的Bph DOX,其中一个大肠杆菌克隆表现出对苯、甲苯和烷基苯极强的降解能力。这种酶中的4个氨基酸残基(H255Q,V2581,G268H和F277y)从KF707改变成LB400酶。随后的定点突变说明这些氨基酸决定单环芳香烃的降解。
Barriault D等以来源于伯克霍尔德氏菌菌株LB400、睾丸酮丛毛单胞菌B-356(Comanonas tostosteroni B-356)和圆红球菌P6(Rhodococcus globerulus p6)的bphA基因的关键片段作为同源基因进行家族DNA改组,以儿茶酚代谢物作为检测手段所得到的几种BPhA突变体表现出更强的降解PCBs的能力,也显示出对2,2'-,3,3'-和4,4'-一氯二苯的更强的降解活性。
(4)未能培养微生物生物降解信息资源的开发利用
目前得到充分利用的生物降解信息资源主要来自可培养微生物,但未能培养微生物也含有丰富的降解信息资源,这部分资源的开发利用对生物降解的强化可以起重要作用。
一般认为环境样品中的微生物中仅有3%可被培养,而其余的97%未能培养,前者称为可培养微生物(culturable microorganisms),而后者称为未能培养微生物(unculturable microorganisms),也被称为活的未能培养微生物(viable butnonculturablemicroorganisms)。从遗传学上说未能培养微生物中的一部分有与可培养微生物同样或相似的系统发育水平,但因对其生理、营养需求知识缺乏而不能应用现行培养技术培养出来,或因它们某些生理学原因抗拒现行培养方法而不能培养。而另一部分则代表新的完全与可培养微生物不同的新谱系(novel lineages),它们根本不能用标准的方法加以培养。
现在所进行的研究工作说明污染环境中不但可培养微生物携带有降解污染物的遗传信息,未能培养微生物也携带有贡献于生物降解的遗传信息。
传统上的微生物多样性依赖于微生物培养技术,而现在评估多样性的分子生物学方法已经跨越传统的培养的方法,成功地用于评价和解释各种环境上的分类多样性,这也包括各种污染地点的多样性(表8-1)。最近,分子技术已发展了特征化和监测各种环境的功能多样性,图8-3是其中的流程图表。Abed RM等应用这种方法研究受石油烃严重污染地在污染物降解后的微生物聚丛及其群落的变化,受污染地生态系统微生物多样性的分析表明这种系统多样性包括培养和未能培养微生物的多样性,两者的多样性都贡献于生物降解。Marchesi J R和Weightman AJ比较研究可培养的α-卤代羧酸(α-halocarboxylic acid)降解菌的脱卤酸基因库和环境样品的总基因组(metagenome)(宏基因组)直接分离的核酸(direct isolation of nucleic acids),结果表明革兰氏阳性细菌占有总多样性的显著部分,然而由于培养的障碍,建立在富集技术基础上的研究至今尚未报告有革兰氏阳性的α-卤代羧酸降解菌。Mac-Naughton S J等用磷脂脂肪酸和变性梯度凝胶电泳分析方法对照研究了石油污染地和未受污染地的微生物群体差异,发现对照点缺乏α-变形细菌(α-proteobacteria),而实验点却含有范围广泛的该类细菌。这说明有受污染地中未能培养的微生物参与石油烃的生物降解。
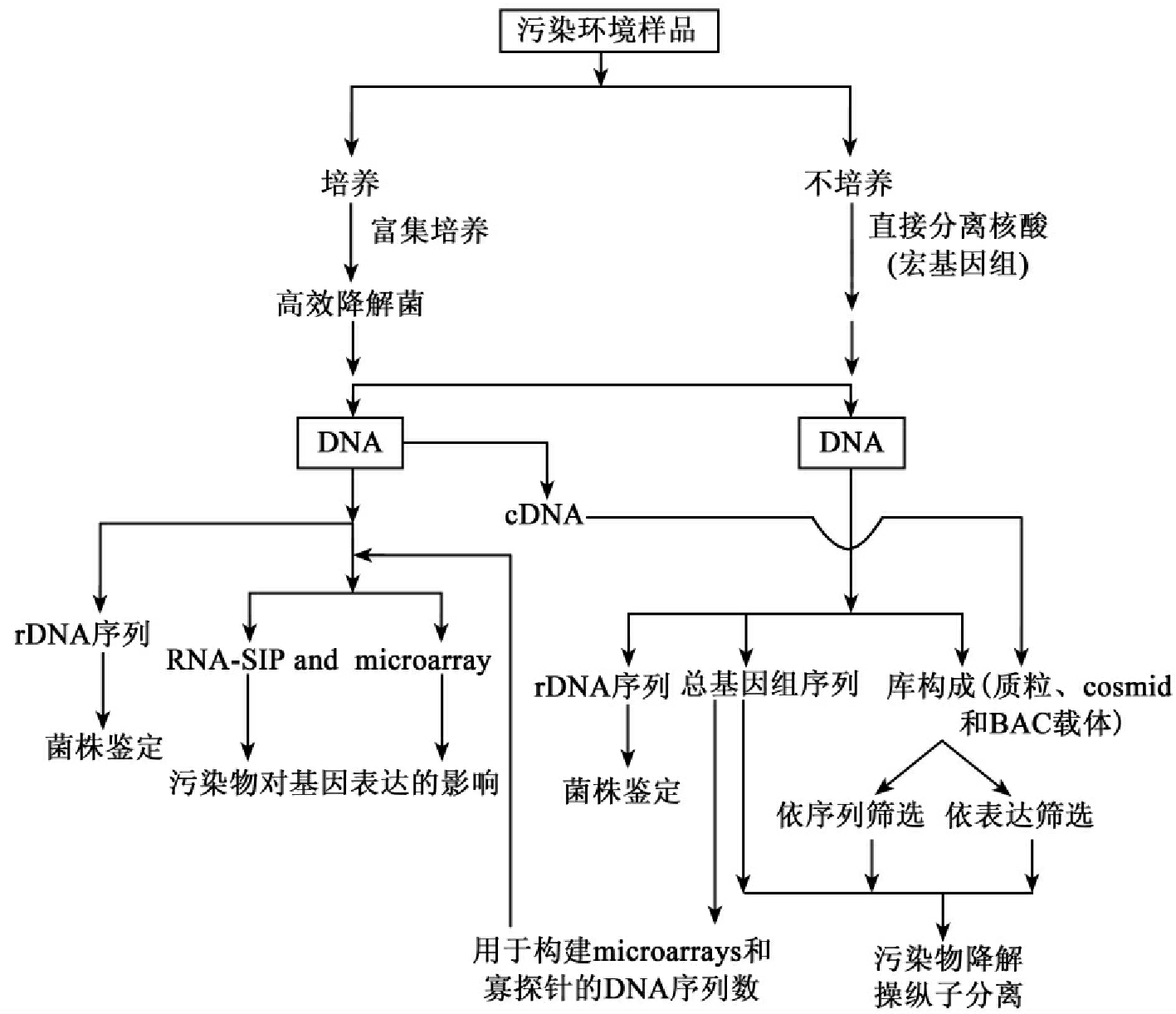
图8-3 最新生物技术研究污染环境的分类和降解多样性
注:BAC:细菌的人工染色体;FISH:荧光原位杂交;RNA-SIP:RNA稳定同位素探针。
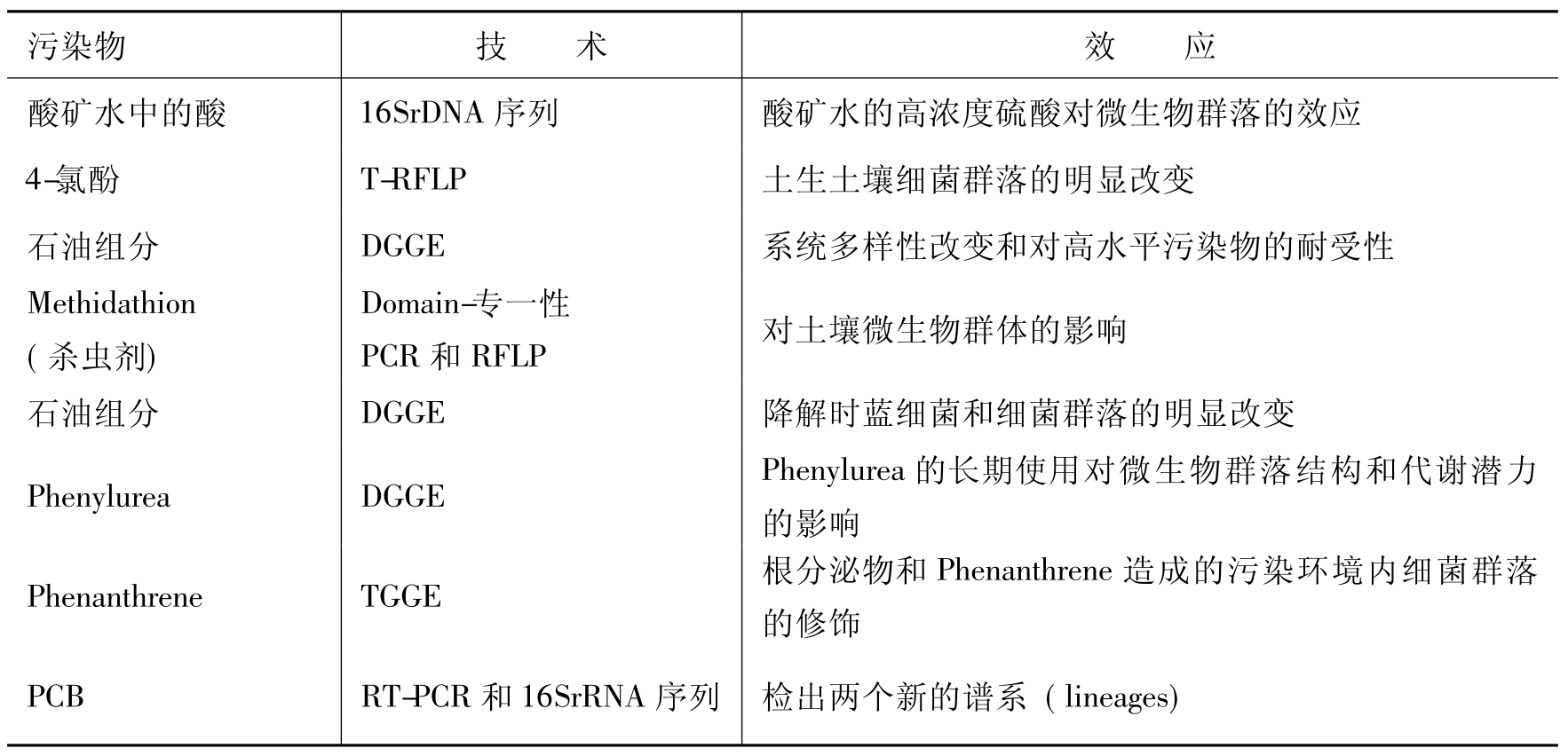
表8-1 分子生物学指标指示污染地中污染物对群落结构的效应
从环境样品中提取的总基因组(基因片段)是巨大的可利用资源,它们可以被克隆到细菌的人工染色体,fosmids或cosmid载体,进行序列基础和表达基础的筛选,或者对整体的基因组序列(以功能序列为基础)直接进行筛选,从而分离和筛选出降解污染物的操纵子。同时利用这种资源可以建立宏基因组的巨大数据库,进一步推动宏基因组的开发利用,这将促进更加有效的“设计型生物催化,促进生物修复。”
设计超级生物催化剂能力的策略最需要的是增大降解生物在环境条件下的可靠表达,从这个角度说理解异生物源化合物和生物之间相互作用的研究以及微生物在环境的归宿、存活和活性,同时还和生物化学和遗传工程研究相交织在一起,这样一种交互(cross feeding)将提供成功干涉环境过程的基础。
第三节 微生物的降解质粒
一、微生物降解遗传信息的分布
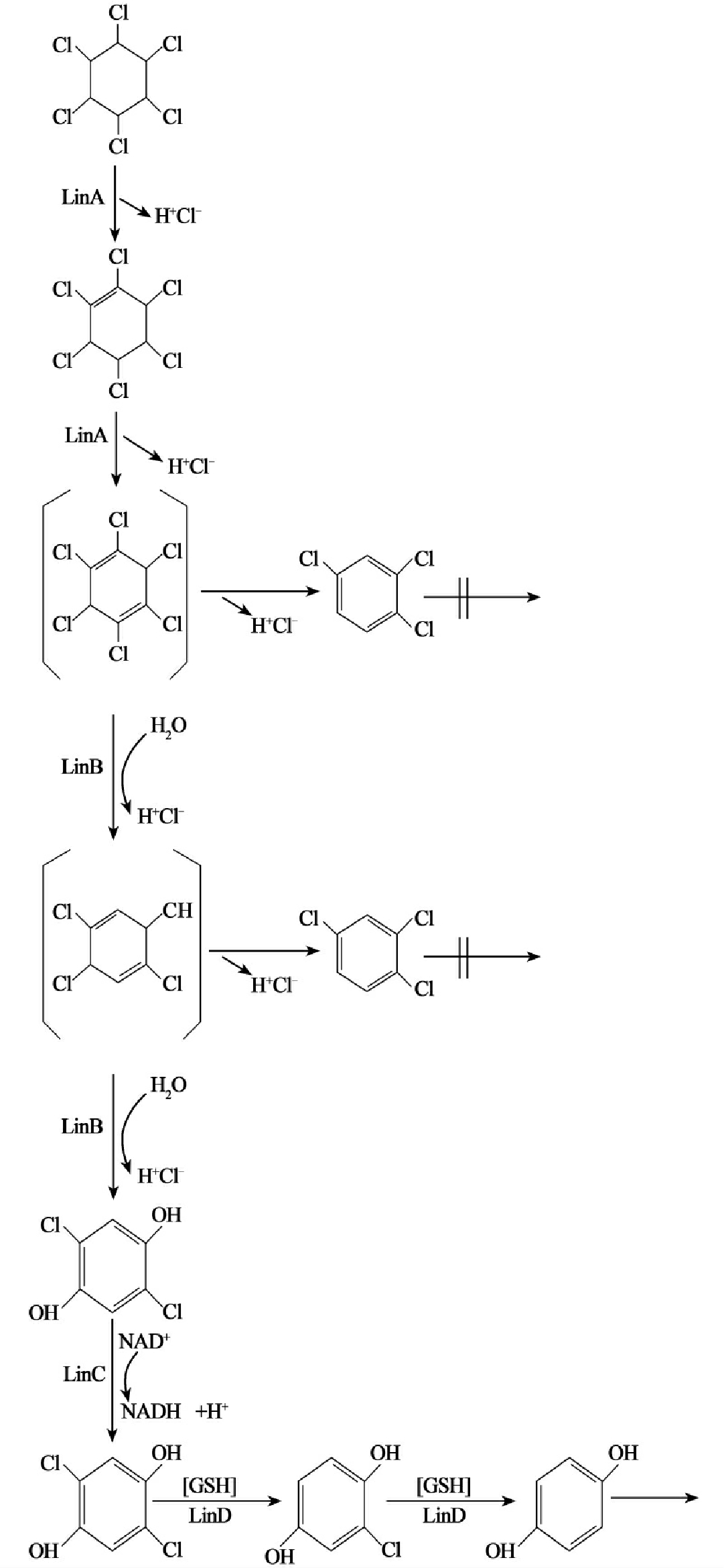
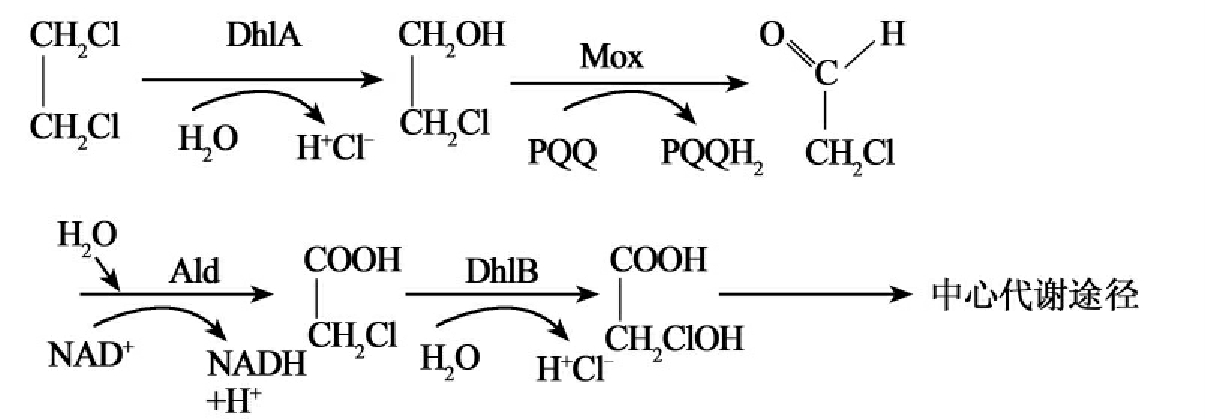
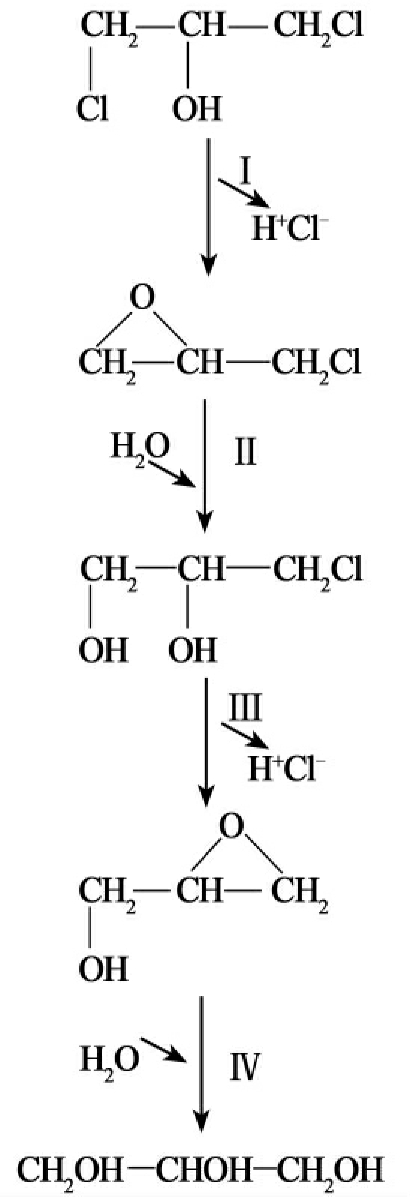
染色体DNA作为细胞中的主要遗传因子,携带有在所有生长条件下所必需的基因,这些基因被称之为“持家基因”,而质粒所含的基因对宿主细胞一般是非必需的,只是在某些特殊条件下,质粒能赋予宿主细胞以特殊的机能,从而使宿主得到生长优势。降解性质粒携带有能降解某些化合物(如芳香族化合物、农药等)的酶基因,这为微生物扩大基质范围,利用难降解化合物(非生物源化合物)生长成为可能。微生物对有机污染物的降解利用拓展了微生物原有的功能,超出染色体上基因组的作用范围,而最终又使微生物从中得到能量和生长物质。这正是原来染色体上基因的正常功能。由此我们看到有机污染物的生物降解过程中的酶系从总体上是由染色体和质粒共同编码的,微生物降解污染物的遗传信息是由染色体和质粒共同携带的。微生物降解有机污染物,特别是降解那些难降解有机污染物的途径复杂多样,降解基因的进化过程十分曲折,因此其降解遗传信息(降解基因)在染色体、质粒中的分布也是多种多样的。从现在的研究结果看,一般有三种情况:①对易降解的有机污染物其降解酶大多是由位于染色体上的基因编码的。②对难以降解的有机污染物,一般前半部分的降解由质粒上的基因编码酶催化,并产生易于矿化利用的中间代谢产物。其后的降解(后半部分降解)则是由染色体上的基因编码酶进行的。③难降解化合物的前半部降解有时也会由质粒和染色体的基因编码酶共同完成,而后半部分的降解过程则由染色体上基因编码的酶进行。1,2-二氯乙烷的降解就是这样(图8-4)。
二、质粒与降解质粒
1.质粒
质粒(plasmid)是细胞中独立于染色体,能进行自主复制的细胞质遗传因子,主要存在于各种微生物细胞中。质粒通常以共价闭合环状(covalently closed circle)的超螺旋双链DNA分子存在于细胞中。这种构型称为ccc型,此外还有oc型(open circular form)和L型(linear form)。近年来还在疏螺旋体、链霉菌和酵母菌中发现线型双链DNA和RNA质粒。质粒分子的大小范围从1kb左右到1000kb。

图8-4 自养白色杆菌降解1,2-二氯乙烷的途径,降解基因分别定位在质粒和染色体上
它们最基本的特性是能寄生在宿主细胞中,并和宿主细胞进行同步复制,在细胞分裂时,能保持恒定地传给子代细胞。一般来说,质粒的存在对宿主细胞并不是必需的,没有它细胞照样生存。在某些特殊情况下,质粒能赋予宿主细胞特殊的机能,从而使宿主细胞得到生长的优势。例如,抗药性质粒和降解质粒就能使宿主细胞在具有相应药物或化学毒物的环境中生存和发展。由于质粒携带细胞编码多种遗传性状的基因,它们是细菌进化的一个重要因子,因为要使宿主细胞迅速适应改变的环境条件,基因就得扩增,从而进行种内和种间的转移。
根据质粒所编码的功能和赋予宿主的表型效应,质粒被分为不同的类型。
(1)致育因子(fertility factor,F因子)
致育因子又称F质粒,F质粒在大肠杆菌的接合作用(conjugation)中起主要作用。
(2)抗性因子(resistance factor,R因子)
抗性因子又称为R质粒,主要包括抗药性和抗重金属。带有抗药性因子的细菌对多种抗生素和其他药物呈现抗性。如R1质粒(94kb)可使宿主具有对氯霉素、磺胺等的抗药性,另外一些R质粒能抗碲、砷、汞、钴、银、镉的毒性。
(3)col质粒
col质粒因首先见于大肠杆菌而得名,其含有编码大肠菌素的基因,大肠菌素是一种细菌蛋白,只杀死近缘且不含col质粒的菌株,而宿主不受其产生的细菌素影响。
(4)毒性质粒
许多致病菌的致病性是由其所携带的质粒引起的,质粒具有编码毒素的基因。例如苏云金杆菌含有编码δ内毒素(伴孢晶体)的质粒,此外Ti质粒也属于此类。
(5)代谢质粒
代谢质粒含能编码降解酶的基因,也称为降解性质粒。另外一些能编码固氮功能的质粒也属代谢质粒范围。
(6)隐秘质粒
隐秘质粒是不显示任何表型效应的质粒,但它们的存在可以通过物理的方法检测。
质粒的功能极为多样,上述的质粒分类并不能包括全部的质粒表型,表8-2列出了细菌质粒赋予细胞表型的例子。
表8-2 细菌质粒赋予细胞表型的例子

续表
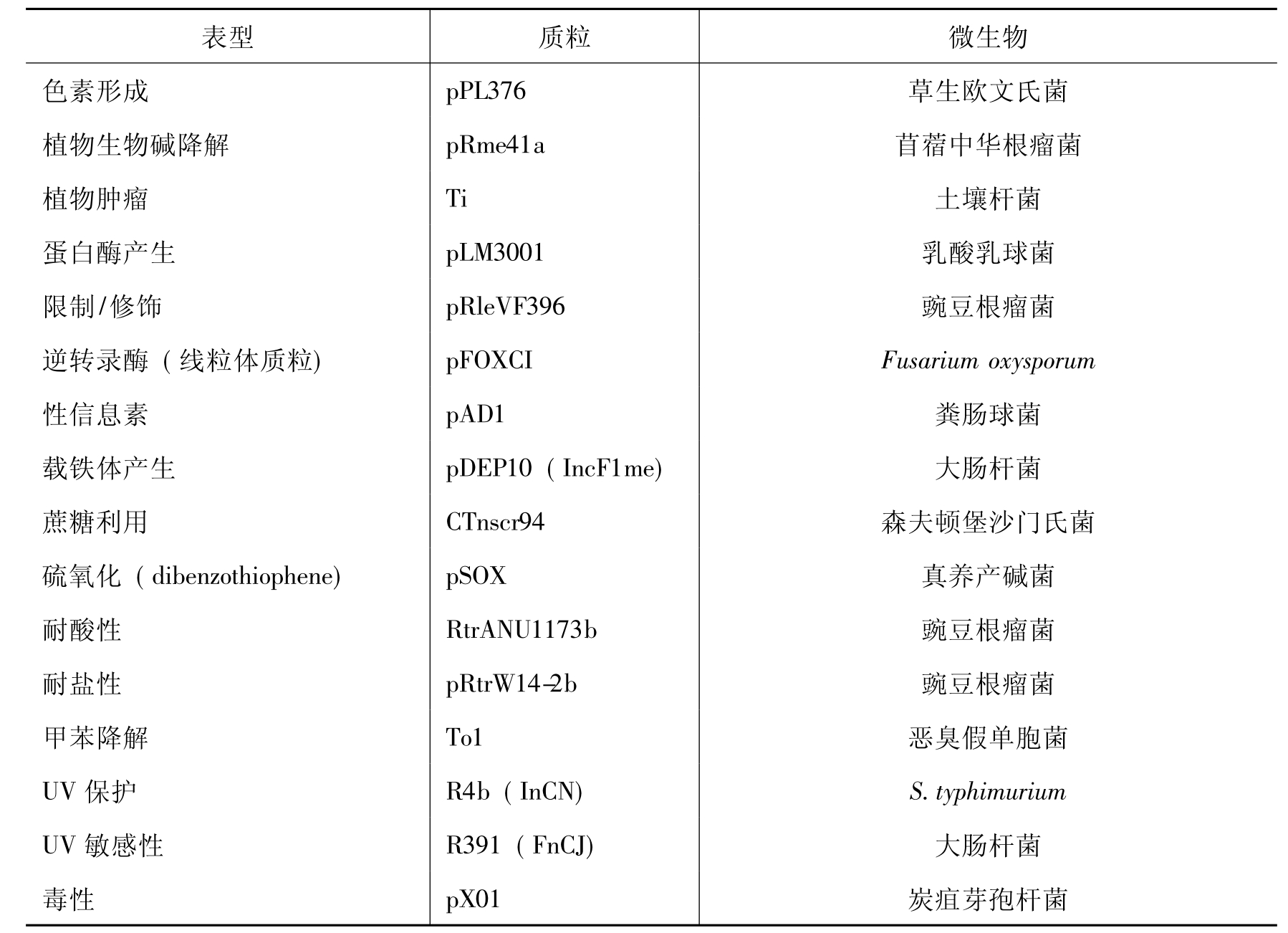
2.降解性质粒
降解性质粒是质粒中特别重要的一类,许多难降解化合物的降解酶类是由质粒上的基因编码的,这类质粒被称为降解性质粒(catabolic plasmid)。当然质粒编码的酶也可参与生物物质的降解。降解性质粒的分子量较大,一般为50×106~200×106U。细菌中的降解性质粒和分离细菌所处环境污染程度密切相关,从污染地分离到的细菌50%以上含有质粒,与从清洁区分离的细菌质粒相比,不但数量多,其体积也大(信息量大)。
自从1972年美国学者Chakrabarty发现降解水杨酸盐的SAL质粒以来,研究人员相继在假单胞菌、黄杆菌、气单胞菌等菌株中发现许多降解性质粒。到目前为止,从自然界分离的菌株中发现的天然降解性质粒共约有50多种。其中666、氯苯、木质素、烷基苯磺酸、SDS、P-羟基苯甲酸、3,5-二甲苯酚、2,6-二氯甲苯等化合物的降解,都是由降解性质粒控制的。被广泛深入研究的部分质粒列于表8-3。
表8-3 部分降解性质粒及其降解底物
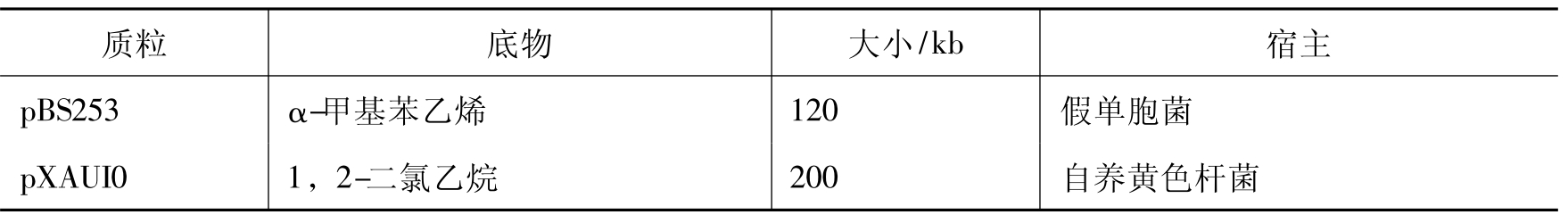
续表
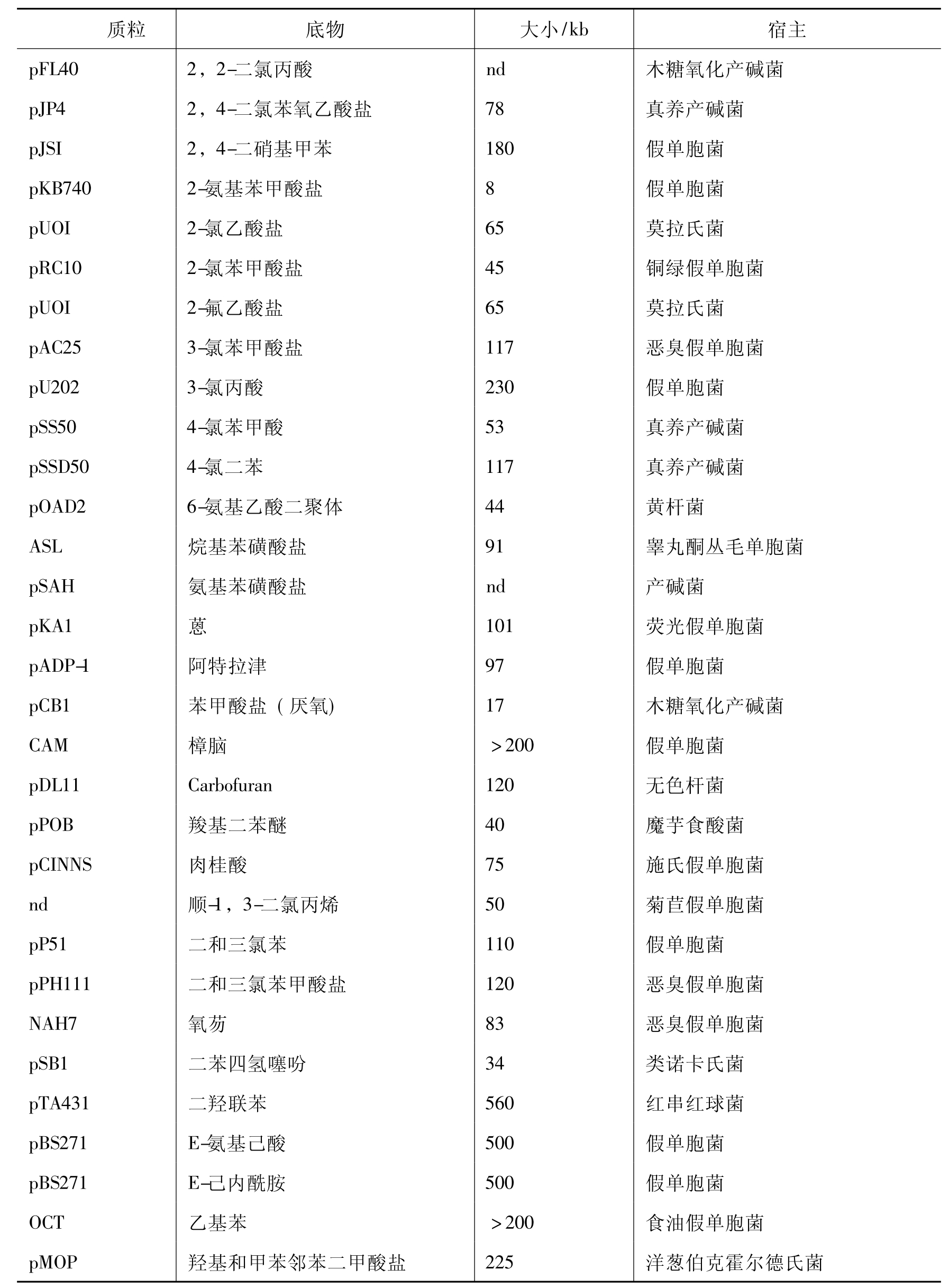
续表
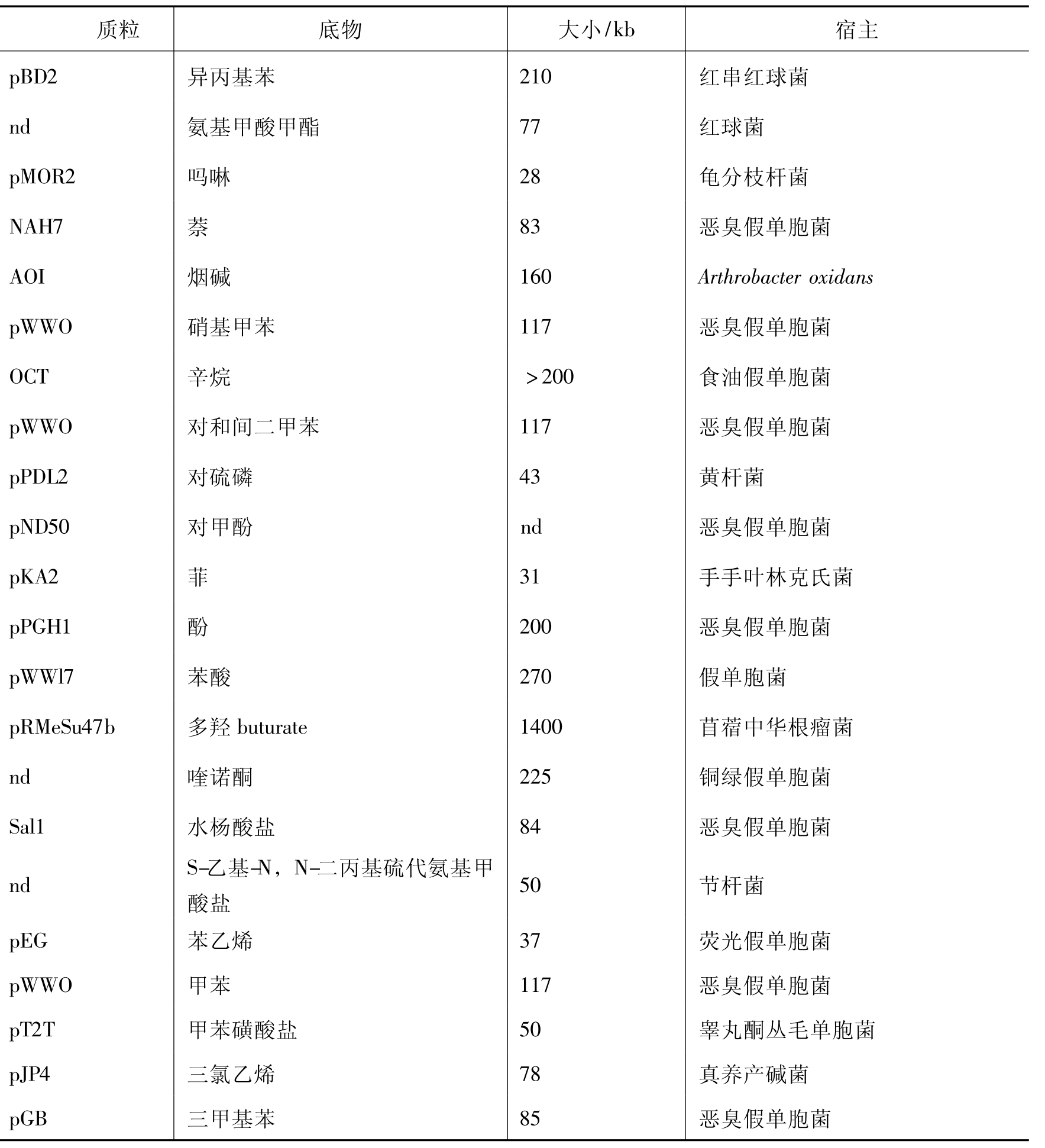
注:nd表示未测定。质粒命名:每一个新分离到的质粒都给予一个编号,一般是小写字母p代表质粒,大写字母表示分离命名实验室的缩写,阿拉伯数字表示序号,如pXy1234。降解质粒也有以降解基质缩写(大写)命名的,如OCT。
三、降解质粒与降解过程
部分带有降解质粒的细菌对其底物的降解途径以及质粒编码的酶在降解中的作用被深入进行了研究。位于质粒上的降解基因编码的酶催化降解过程的前期反应,降解过程的中间产物又可被染色体上基因编码的酶进一步降解。从这些降解过程中,我们可以看到微生物降解质粒在生物降解中的重要作用。
质粒的降解底物非常多样,大多数污染物特别是难降解污染物的生物降解都有质粒的参与和起作用。下面以萘、甲苯和2,4-D的降解为例说明质粒的重要作用。
1.萘的降解
恶臭假单胞菌PPG7的NAH7质粒中基因编码酶降解萘途径如图8-5。萘通过不同的途径进入环境,从环境中已分离出许多降解萘的微生物,携带NAH7质粒的恶臭假单胞菌PPG7对萘的降解性具有代表性。质粒中的降解基因组成两个操纵子。上游操纵子编码把萘转化成水杨酸盐的酶,而下游操纵子编码的酶把水杨酸盐转化成TCA循环的中间代谢产物。多组分的萘加双氧酶催化上游途径中的第一步反应,把分子氧插入1,2位双键的两边。萘加双氧酶由4个亚基组成,酶由nahA基因簇中nahAa、Ab、Ac和Ad分别编码。反应产物是顺-dihydroldiol萘。这种化合物的羟化环被dihydrodiol脱氢酶(NahB)再芳香化(rearomatized),导致失去两个质子。1,2-二羟萘双加氧酶(Nahc)加入分子氧断开羟化环的双键。这种环断开产物自发异构化成为2-羟基苯并吡喃-2-羟酸盐。2-羟基苯并吡喃-2-羧酸盐异构酶(NahD)催化二次异构化得到2-羟基苯基丙酮酸盐。这种分子被具有水解酶和醛缩酶功能的同一种酶(NahE)水解断开烷烃侧链,而后再被氧化,反应产物是水杨醛,水杨醛被氧化酶(NahF)进一步氧成水杨酸盐。
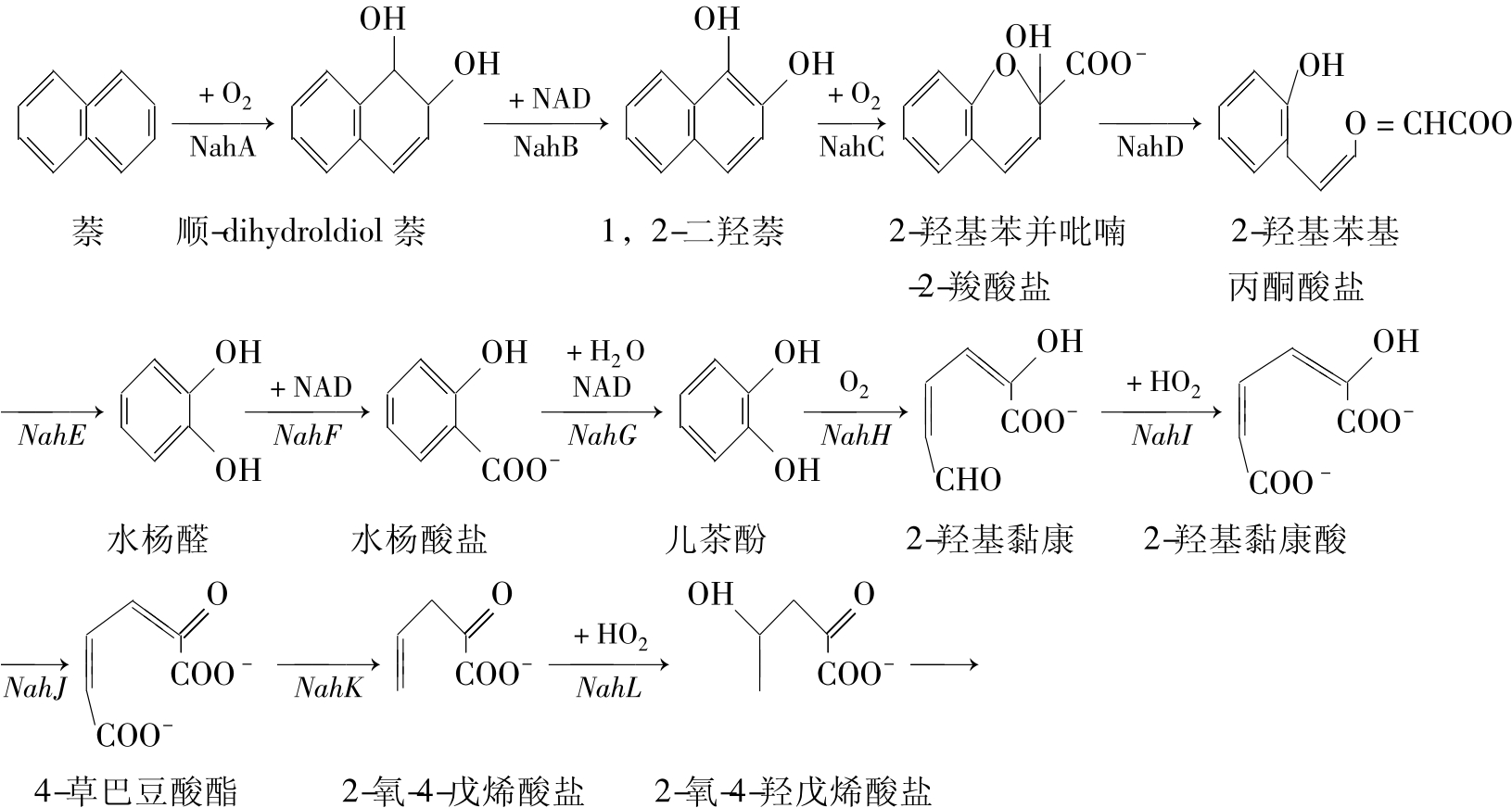
图8-5 NAH7质粒上基因控制的萘降解途径
下游途径的第一步是水杨酸盐水解酶(NahG)催化水杨酸盐单加氧氧化形成儿茶酚,儿茶酚氧化的第一步是环的间位断开,nahH编码的儿茶酚-2,2-双加氧酶催化这一反应,把分子氧加到邻接一个羟基的双键两端。这种间位断开不同于邻位断开(邻位在2,4-D的降解中见到)。产物2-羟基黏康半醛能被进一步代谢,通过水解去除aldehyde基团(NahN)得到2-氧-4-戊烯酸盐。另一途径是aldehyde首先被氧化成2-羟基黏康酸盐(NahI),而后又被tautomerized成4-草巴豆酸酯(NahJ),其后在草巴豆酸酯脱羟酶(NahK)作用下也得到2-氧-4-羟戊烯酸盐。其后在2-氧-4-戊烯酸盐水解酶(NahL)水解作用下产生2-氧-4-羟戊烯酸盐。最后这种分子被染色体基因编码的醛缩酶打断得到丙酮酸和醛,两者都进入TCA循环。
在本降解过程中编码间位断开途径的基因是水杨酸操纵子的部分,而实际上儿茶酚的降解情况是复杂多样的,一般能降解芳香化合物的微生物都能代谢儿茶酚。编码儿茶酚降解的基因已被定位在某些生物的染色体上,有的定位在质粒上,而某些生物可以把拷贝同时定位在染色体和质粒上。
2.甲苯的降解
pWWO(TOL)质粒不但能降解甲苯,也能降解m-和P-二甲苯(m-和P-xylene)和其他苯衍生物。质粒的结构如图8-6所示,编码降解酶的操纵子被命名为xy1。基因由两个操纵子组成,两个操纵子涉及上游(upper)和下游(lower)间位裂解途径。它们编码的酶列
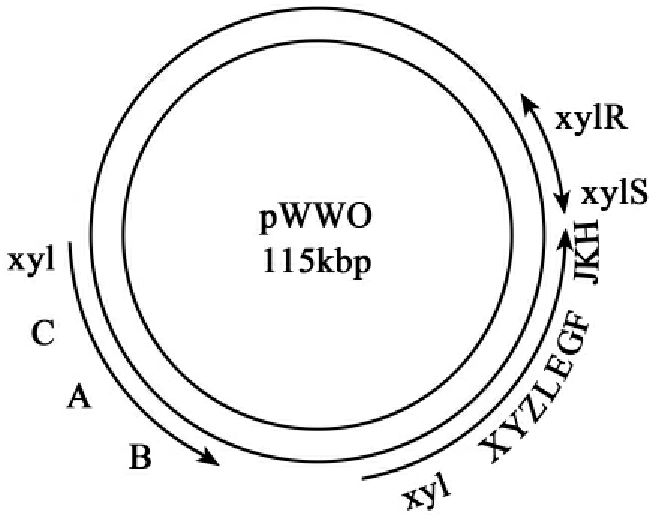
图8-6 pWWO(TOL)质粒图
如表8-4所示,xy1CAB编码的酶把甲苯降解成苯甲酸,而xylXYZIEGFJKIH编码的酶把苯甲酸降解成乙醛和丙酮酸。携带pWWO质粒的恶臭假单胞菌mt-2菌株降解甲苯途径如图8-7所示。

图8-7 pWWO质粒编码酶主导的甲苯降解上游途径
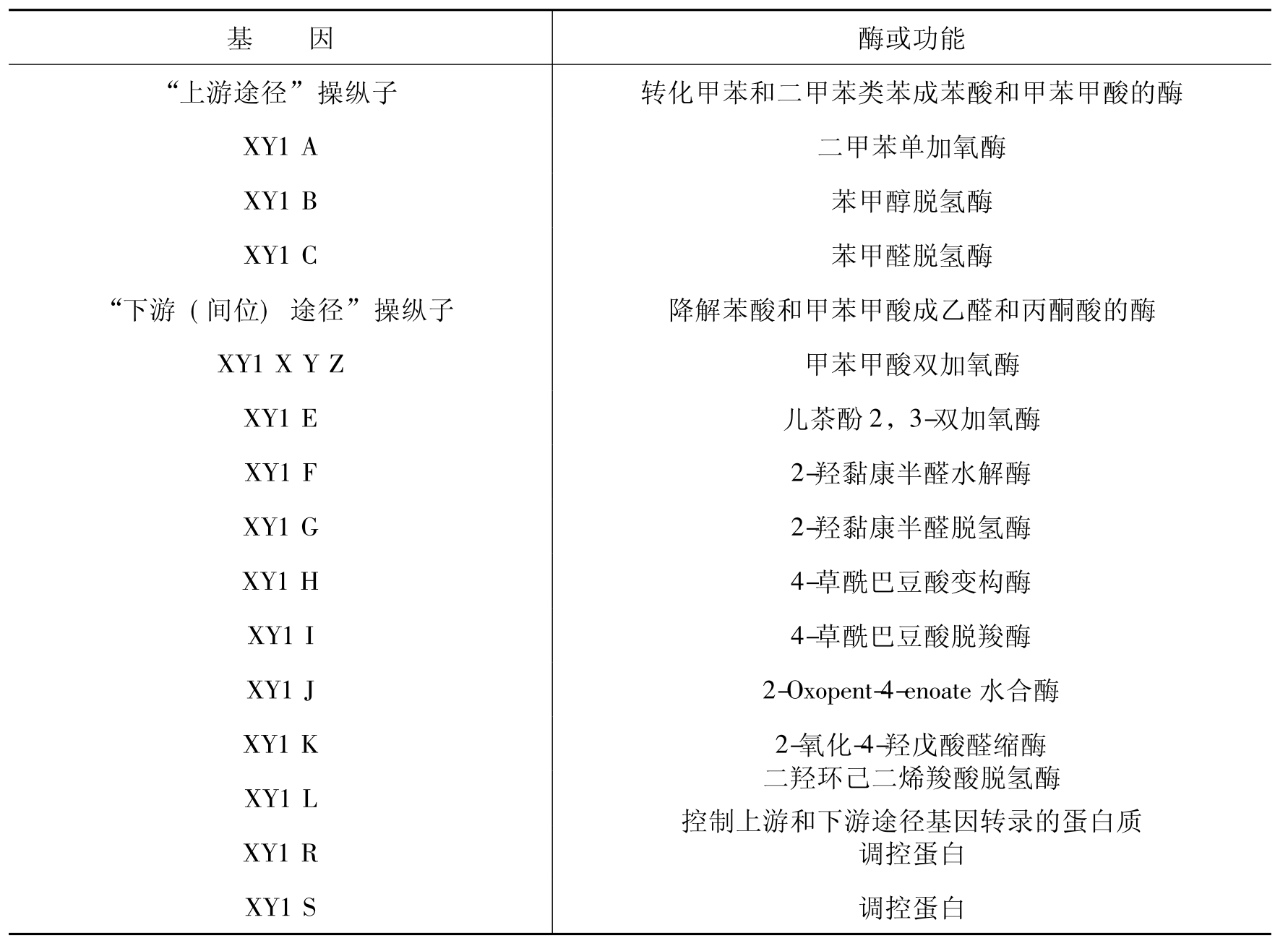
表8-4 pWWO(TOL)质粒基因编码的酶和调控蛋白
3.2,4-D的降解
能降解2,4-D的真养产碱菌菌株JMP134(A.eutrophus JMP134)的2,4-D降解质粒PJP4是被广泛深入研究的具有代表性的质粒。这种质粒广宿主范围(broud host-range plasmid),大小78kb,能自主转移(self-transmissible)。6个结构基因(tfdA、tfdB、tfdC、tfdD、tfdE和tfdF)和两个调控基因(tfdR和tfdS)参与降解过程。六个结构基因组成3个操纵子fdA、tfdB和tfdCDEF,编码6种降解酶,把2,4-D降解转化成β-氧代己二酸。这个最后产物成为染色体编码的己二酸途径的底物,最后得到琥珀酸和乙酸盐进入TCA循环。质粒编码酶对2,4-D的降解途径如图8-8所示。
4.质粒的水平转移
上述的例子说明质粒编码的酶参与单一一步到复杂的多步途径,质粒以多种方式使微生物得到代谢能力来开发不能利用的资源。把质粒携带菌株接种到环境中,质粒中的遗传信息还可以转移到土著微生物中。这种类型的遗传交换(称为水平转移)使降解基因具有高水平的同源性,而亲缘关系很疏远的微生物能转化同样的底物。
5.降解基因的同源性与差异
虽然许多降解基因之间有很高的同源性,但DNA的序列和某些更复杂途径的调节控制也存在明显的差异。clc(pAC25质粒)、tcb(PP51质粒)和tfd(质粒pJP4)操纵子的比较研究可以说明,这三种操纵子都编码一种调控蛋白和降解氯代儿茶酚的酶,都有高水平的同源性。然而一定序列的差异已经改变调控蛋白的专一性。clc和tcb以及clc和tfd途径的调控蛋白可以互换,然而tcb和tfd的调控蛋白不能互相替代。这说明这些基因可以有相同的起源,但出现差异,这可能是进化过程和一些其他选择压力所产生的突变积累的结果。
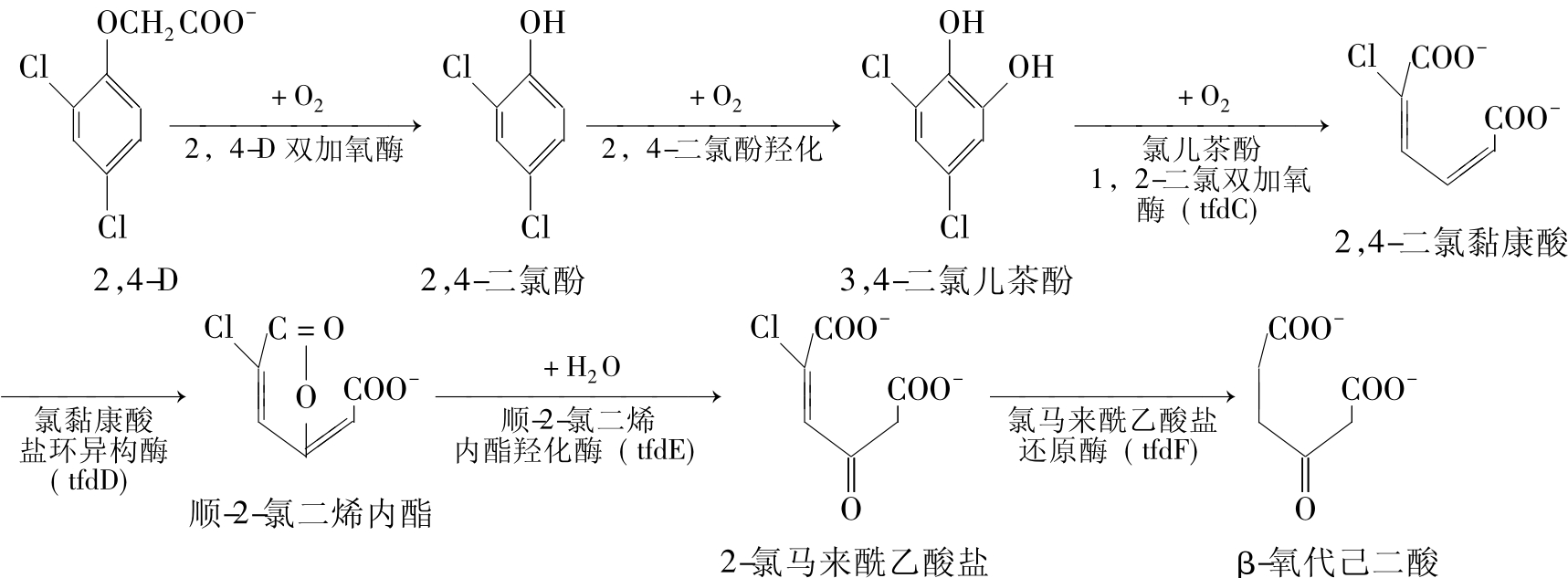
图8-8 真养产碱菌(PJP4质粒)降解2,4-D的途径
四、转录调控
原核生物的基因调控主要发生在转录水平上,这是一种最为经济的调控。功能相关的基因组成操纵子结构,其受同一调节基因和启动子的调控。调节基因通过产生阻遏物或激活物来调节操纵区,从而控制结构基因的功能。质粒上与降解作用相关操纵子的结构基因的转录也和原核生物结构基因一样受到多层次的调控,但操纵子自身的调节基因的调控起重要作用。这些调节基因(如nahR、xylR、xyLS、tfdR、tfdS、DMPR等)编码的LysR、Ntrc、Arac等家族蛋白(family protein)激活或抑制结构基因的转录,这些调节蛋白中最重要的是LysR型蛋白。几种LysR型调控蛋白的例子示于图8-9。
在恶臭假单胞菌PPG7的NAH7质粒中,nahR编码36kU Lys型调控蛋白。这种蛋白质对编码萘降解的降解酶基因的转录作正调控控制。上游途径(nahABCFDE)把萘转化成水杨酸盐,这些基因是低水平组成型表达。下游途径转化水杨酸盐成2-氧-4-羟戊烯酸盐。NahR结合到上游和下游途径操纵子的启动子区域。水杨酸盐是上下游途径基因转录的正调控效应物,当水杨酸盐存在时,NahR构型改变,它能更紧密结合到启动子部分。DNA结合NahR和RNA多聚酶的结合增加频率,成功的转录开始。不像某些LysR型调节子,编码萘降解的基因的转录不受易于被微生物利用的化学物(如琥珀酸盐、葡萄糖或复合的丰富培养基)抑制。萘降解的调控代表一种在分子水平上对整个代谢途径控制的有效方式。这种效率对存在于代谢资源高度有限环境中的细菌群体十分重要。
携带pWWO质粒的恶臭假单胞菌降解甲苯过程由xylR和xylS产生的两种XysR型调控蛋白调控。XylR正调控上游途径,在甲苯存在时,XylR结合到启动子,上游操纵子和xylS基因一般是正调控的低水平组成型转录。由于xylS是下游途径的调控蛋白,两种操纵子被XylR以稍微不同的方式实行调控。XylS一般微弱结合到下游操纵子的启动子。上游途径对甲苯的代谢导致效应物分子的积累(如苯甲酸盐)。在存在效应物时,XylS激活下游途径的转录。如果苯甲酸盐直接作为碳源被提供,XylS也具有独立于XylR的功能,对下游操纵子正转录调控。下游途径也可以被染色体编码的LysR调控蛋白所激活,这种LysR型调控蛋白对染色体编码的苯甲酸盐加双氧酶基因的转录负责。葡萄糖或琥珀酸盐存在时能抑制甲苯的降解。

图8-9 质粒携带降解操纵子的示意图(箭头代表转录方向)
在质粒PJP4起重要作用的2,4-D降解系统中,LysR型调控蛋白被称为tfdS,是由两个同样的基因tfdR和tfdS编码。这种蛋白质正调控tfdA和tfdB两个操纵子的转录,它们编码把2,4-D降解成3,5-二氯儿茶酚的酶,这种蛋白质也调控tfdCDEF和tfdD11。在存在二氯黏康酸盐(TfdD的代谢产物)时,调控蛋白和被调控基因的操纵子的结合受到加强,从而产生转录的正调控。在质粒中还有另外一个tfdD拷贝(称为tfdD11)是从tfdR启动子趋异转录的,这种tfd有更大的产量,另一个基因tfdT在起源上与LysR型调控蛋白相同,tfdT是一个来源于插入事件的残迹(remnant),得到一个无功能的截短蛋白。
五、转座因子
转座因子是细胞中能改变自身位置的一段DNA序列。原核生物中的转座因子有三种类型:插入顺序(insertion sequence,IS)、转座子(transposon,Tn)和某些特殊病毒(如Mu、D108)。IS和Tn有两个重要的共同特征:它们都携带有编码转座酶的基因,该酶是转移位置,即转座所必需的,另一个共同特征是它们的两端都有反向末端重复序列(inverted terminal repeat,ITR)。转座因子的转座可引发多种遗传学效应,主要包括插入突变、产生染色体畸变和基因的移动和重排。
降解质粒中的转座子对降解能力的形成及转移有极其重要的作用,这主要包括作为降解信息的遗传载体,促进降解遗传信息的迁移和重组以及由此而产生的降解能力的进化。
降解质粒中普遍存在转座因子,其和降解操纵子的遗传重排和基因重复有密切关系。位于转座因子上的降解操纵子能转移到受体菌株的质粒或染色体DNA上,这种运动增加了基因水平迁移的潜力。源于质粒PWWO的转座子Tn4651带有完全降解甲苯基因,并带有转座所需的三个基因tnPA、tnPS和tnPT。Tn5280是pP51质粒上的降解转座子。质粒中tcbAB和tcBCDEF能使氯苯降解,其中tcbAB操纵子位Tn5280。插入序列也常见于质粒中,IS6100见于OCT质粒中的alk基因、pXAV质粒中的dhl基因、质粒pAD2的nyl基因中,在最后的质粒中5次出现。这种序列也发现于质粒pNAD2中。质粒pJP4的2,4-D(tfd)降解途径中含有一个插入序列(ISJP4)。ISJP4携带tfdT基因,tfdT基因能产生一种和其他LysR型转录调控物同源的蛋白质。但这种蛋白质没有可见的(discernable)功能,不能激活tfdCDEF操纵子。这是由于ISJP4能干扰tfdT的开放读码,导致产生一种截短的、非功能的TfdT蛋白。
第四节 微生物对异生物源有机物降解能力的遗传进化
在漫长的生物进化中,生物(主要是微生物)对各种各样生物源的有机物的降解能力和这些有机物的存在是相伴产生的。在自然环境中没有任何一种有机物明显过剩,过量积累就很好说明这样的问题。20世纪初叶展开的工业革命,特别是大量的人工合成化合物和原来没有进入自然循环的化合物(如石油中的一些多环芳烃成分)进入自然环境,各种污染物进入环境实际上可以创造出使降解基因进化增殖的生态位。面对这些陌生的外来者,面对原存酶所不认识的底物,暴露在这些新化合物面前的微生物能进化出对这些化合物的降解能力,这就是微生物降解异生物源有机物的遗传进化。
生物进化始终是生物学研究的重要课题,分子生物学的新研究技术及研究成果为生物进化研究提供了新的推动力。同样也为生物降解的遗传进化研究提供了基础。
一、遗传进化证据
异生物源有机物进入自然环境后会不断积累而达到危险的程度,产生无法规避的风险,至今的大量研究说明各种各样的有机污染物除了可被物理、化学、水解和光解外,都在不同程度上可被生物降解。许多以难降解污染物为唯一碳源能源的微生物已经分离出来,例如被公认极难降解的DDT在其使用的短期内人们不能从自然环境中分离到利用这些化合物的微生物,而经过一段时间以后,特别是在20世纪末,已有大量的文献报道,已经分离到许多可以DDT为唯一碳源生长的微生物,这说明微生物在长期的适应过程中进化出降解能力。芳香族化合物及其氯代芳香族化合物种类繁多,其降解过程及降解信息得到充分的研究,研究的结果为生物降解的遗传进化提供了许多确切的证据。这包括:
①已经分离能完全降解利用氯代芳香族化合物的微生物,这些微生物对有机物的降解途径及降解机理都已得到广泛而深入的研究。
②许多不同降解菌株带有相同功能的质粒、降解酶,它们降解过程的代谢调控具有相似性。TOL、NAH和SAL质粒显示出明显的DNA同源性,带有降解氯代芳香族化合物基因的降解质粒pJP4,pAC25,pSS50和pBRC60有一个明显同源质粒骨架,这些骨架决定着复制和转移的功能。氯苯质粒pP51和pJP4降解基因的外侧区域同源。这些结果说明它们共有一个能自我传递的祖先。从美国、澳大利亚和加拿大分离出的2,4-D降解菌,其大部分相应降解基因与真养产碱菌JMP134的pJP4质粒的6个基因,特别是与tfdA有高度的同源性。大部分降解菌拥有类似tfdA、tfdB和tfdC片段的结合,这三种基因的基因探针检测出各种水平的同源性。降解菌中的产碱菌属、伯克霍尔德氏属及红育菌属中的某些菌株和tfdA、tfdC有60%或更高的同源性。
③现在实验室研究利用各种分子生物学技术所组建的新的降解菌株实际上可以认为是自然环境进化过程的强化和实验室再现。
④芳香类及氯代芳香类化合物降解途径都展现出以中心代谢产物为中心的代谢途径,即中心代谢途径(centralmetabolic pathways),也即各类芳香族和氯代芳香族化合物都先后被转化成儿茶酚和氯代儿茶酚,这两种中心化合物再经间位或邻位裂解而被进一步降解。从中我们可以看到这些化合物的降解存在一条中心途径,可以认为这些化合物的降解进化是围绕这条中心途径经水平扩展、垂直扩展后完成的。
⑤调控系统的同源性。调控氯代芳香化合物降解基因的多为LysR家族的成员,这个家族的每种蛋白质至少有一个亚基是共有的。
二、生物降解遗传进化的主要方式
生物降解遗传进化的主要方式是新降解基因(一个或多个)的产生以及由此产生的对已有的降解途径的扩展和补充,把新化合物的部分降解汇入已有的降解途径中,从而形成特定化合物的完全降解途径。例如2,4-D这种农药是一种异生物源化合物,而其初始降解(降解菌为真养产碱菌JMP134)产物2,4-二氯酚是一种自然的化合物,而这种化合物的降解途径早已存在,因此2,4-二氯酚降解菌仅需获得2,4-D降解第一步的基因2,4-D-双加氧酶基因,就可以获得完全降解2,4-D的能力。农药甲基对硫磷的降解也类似,其初始降解第一步水解的产物是P-硝基酚,而能降解这种化合物的细菌广泛存在于土壤和表水中,因此它们仅需进化出甲基对硫磷水解基因即能完全降解这种化合物。当然除了获得一个新降解基因就能完成整个降解途径外,也会有获得多个降解基因才能完成降解的例子。
获得一个新降解基因就可获得降解能力的细菌要比获得多个基因才能完成降解的细菌有更快的降解能力进化速度。2,4-D的降解属于前者的例子,而阿特拉津(atrazine)属于后者,其降解过程中开始的多步降解产物是异生物源化合物。2,4-D在20世纪50年代开始使用,而在20世纪60年代后期就分离到能以其为唯一碳源和能源的细菌,而阿特拉津在1959年就开始使用,而直到1993年后才分离到完全降解的这种农药的细菌。
三、遗传进化的机理
我们现在的实验手段还难以重现微生物生物降解的进化过程,但我们可以从现代分子生物学技术改造微生物成果,并结合对从自然环境中分离到的降解微生物的遗传分析来推断生物降解的遗传进化,阐明推动生物降解遗传化的基本机理。大量的实验研究和理论分析表明从根本上说生物降解的进化可以归结为微生物群落中新的降解基因的出现和基因的相互作用,而最终在一个微生物群落中出现降解特定有机物的全部基因。目前的研究表明基因突变、接合作用、转化、转座造成的新降解基因以及基因转移和重组使微生物进化出新的降解能力是生物降解遗传进化的主要机理。许多代谢活性的实验进化研究说明基因转移和重组对宿主细胞适应新化合物的重要性。通过基因转移和重组能克服新基质降解自然途径的生物化学障碍。基因转移可以产生更广谱的酶替代窄谱的专一性酶而产生代谢途径的水平扩展。也可以提供新的非关键性酶(peripheral enzymes)来介导基质到已存在的降解途径中而产生垂直扩展。
基因突变是基因内部遗传结构或DNA序列的改变,包括一对或少数几对碱基的缺失、插入或置换而导致的遗传变化。单一位点突变(single-site matation)可以在微生物基因中连续发生,从而会改变基因的结构或造成损伤,在DNA复制过程中出现差错。微生物在DNA复制中能对损伤进行修复,包括光复活修复、切除修复、重组修复和SOS修复。但在SOS修复中也产生一种错误倾向(error-prone)的SOS修复,这种修复识别碱基的精确度低,因此容易造成复制的差错,这是一种以提高突变率来换取生命存活的修复。基因突变和修复过程中的差错都可以产生相应的突变,在环境压力存在的条件下,环境压力的选择会积累DNA的进化。有研究证明单一位点突变能改变酶的底物的专一性或效应物的专一性。PWWO质粒中基因编码的儿茶酚2,3-双加氧酶因酶的单一氨基酸被取代而使底物范围扩展到4-乙基儿茶酚。
接合作用是通过细胞与细胞的直接接触而产生的遗传信息的转移和重组过程。
遗传转化是指同源或异源DNA分子(质粒和染色体DNA)被自然或人工感受态细胞摄取,并得到表达的水平方向的基因转移过程。实验研究表明恶臭假单胞菌mt-2转移TOL质粒PWWO到假单胞菌B13菌株,使生物降解能力范围从3-氯苯酸盐水平扩展到4-氯苯酸盐和3,5-二氯苯酸盐。这种转移提供B13菌株以TOL质粒的xy1 XYZ(甲苯甲酸盐双加氧酶),这种酶的底物范围比原菌株B13的氯苯酸盐加双氧酶更广。质粒转移已在微宇宙实验中得到证实,编码3-氯代苯酸盐降解酶的产碱菌菌株BR60的降解质粒pBRC60被转移到土生受体菌并表达。
转座是转座子或插入序列插入同一DNA分子的新的位点或插入另一DNA分子的过程。基因片段的插入可以导致基因转移,实现DNA片段的重排,使沉默基因激活或活跃基因纯化。插入片段对不常见化合物的适应性和降解潜力有重要作用。研究证明洋葱伯克霍尔德氏菌至少带有9种不同的插入片段,它们在基因组中存在1~13个拷贝。插入片段(IS931)在chq基因座(chq gene locus)周围,对2,4,5-T降解起作用。这种片段不来源于这种菌,这说明IS931或这种降解基因的部分是从其他生物中获得。假单胞菌菌株P51的氯代苯加双酶基因的两侧有两种等同插入片段IS1066和IS1067。IS1066、氯代苯加双氧酶基因和IS1067共同组成的称为Tn5280的复合片段也是一个功能转座子,能随机插到基因组。IS片段的另一个重要作用是对沉默基因的激活。IS片段的一端常常含有类似启动子(promoter like)序列,其能激活IS片段外基因的表达。在洋葱伯克霍尔德氏菌中,IS406和IS407的插入能导致lacZ的激活。IS931和IS932也能激活邻近基因的表达。人工构建的菌株可以重现自然条件下所发生的适应进化。例如氯代苯完全降解的途径需要广底物专一性(broadsubstrate-specificity)的苯双加氧酶、广底物专一性的苯glycol脱氢酶,同时要有修饰邻位裂解途径。恶臭假单胞菌F1含有编码广底物专一性的苯加双氧酶和苯glycol脱氢酶的基因,假单胞菌B13的质粒带有裂解途径,通过这两个菌株的融合所得到的转化接合子(transconjugants)能完全代谢氯代苯。而从污染环境中,分离到一株假单胞菌P51也说明这种情况。这个菌株的质粒含有两个降解操纵子,一个编码修饰的邻位裂解途径,而另一个编码氯苯加双氧酶和氯苯glycol脱氢酶。氯代苯加双氧酶基因簇位于可转移的片段上,这说明现在的pP51质粒是氯代苯加双氧酶转座子转座到老pP51质粒上形成的,老的质粒仅含有修饰邻位裂解途径基因。
第五节 微生物的降解反应、降解途径、降解速率及降解指示系统
本节讨论生物降解中的几个基本问题:降解反应、降解途径、降解速率、归宿等基本问题。
一、降解反应
一般来说自然有机物的微生物代谢和有机污染物的生物降解在本质上是一样的,发生在前者的化学过程也见于各种污染物的降解过程中,主要包括氧化反应、还原反应、水解反应和聚合反应。
1.氧化反应
氧化反应是许多有机污染物生物降解开始的第一步,是最重要的反应,特别是许多烷烃降解中的重要反应。羟化作用(hydroxylation)是最一般的氧化反应。例如芳香环氧化成儿茶酚的过程就是羟化作用的结果(图8-10)。形成羟基会增加化合物的极性,从而提高化合物的水溶性,有助于提高生物降解性。催化这种反应的酶包括羟化酶和混合功能氧化酶。另一类重要的氧化反应是N-脱烃作用(N-dealklation),也常是污染物降解的第一步,特别是alkyl取代的农药(烷基取代基农药),N-脱烃作用可被混合功能氧化酶所催化(图8-11)。其他的氧化反应还包括氧化脱氢、氧化脱氯、脱羧作用、β-氧化、醚键的水解、环氧化作用、硫氧化作用(sulfoxidation)及芳香环、杂环的断裂。有些化合物经氧化反应后仍保持原底物的生物活性,如阿特拉津降解的第一步是氧化反应,其氧化反应的产物仍保持作为农药的生物活性。
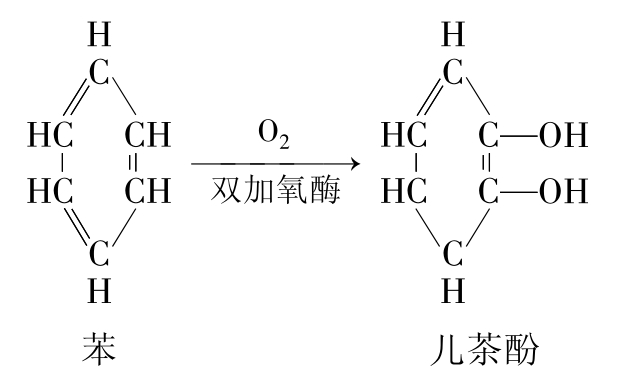
图8-10 苯氧化成儿茶酚的过程
2.还原反应
很多有机污染物在厌氧生物降解时发生还原反应(好氧条件下也有还原反应)。例如还原脱卤,酮(ketones)还原成醇(alchohols),亚砜(sulfoxide)还原成硫化物(sulfide)。在还原反应中,还原脱氯尤为重要,还原脱氯是许多化合物(包括有机氯农药、烷基溶剂、烷基卤化物)的一种重要降解方式。这些化合物包括许多毒性最强,最难降解的污染物。脱氯以后可降低毒性,提高可降解性。DDT加氢转化成DDD就是一个烷基还原脱氯的例子。催化还原脱氯的催化剂是存在于细菌中的过渡金属复合物。
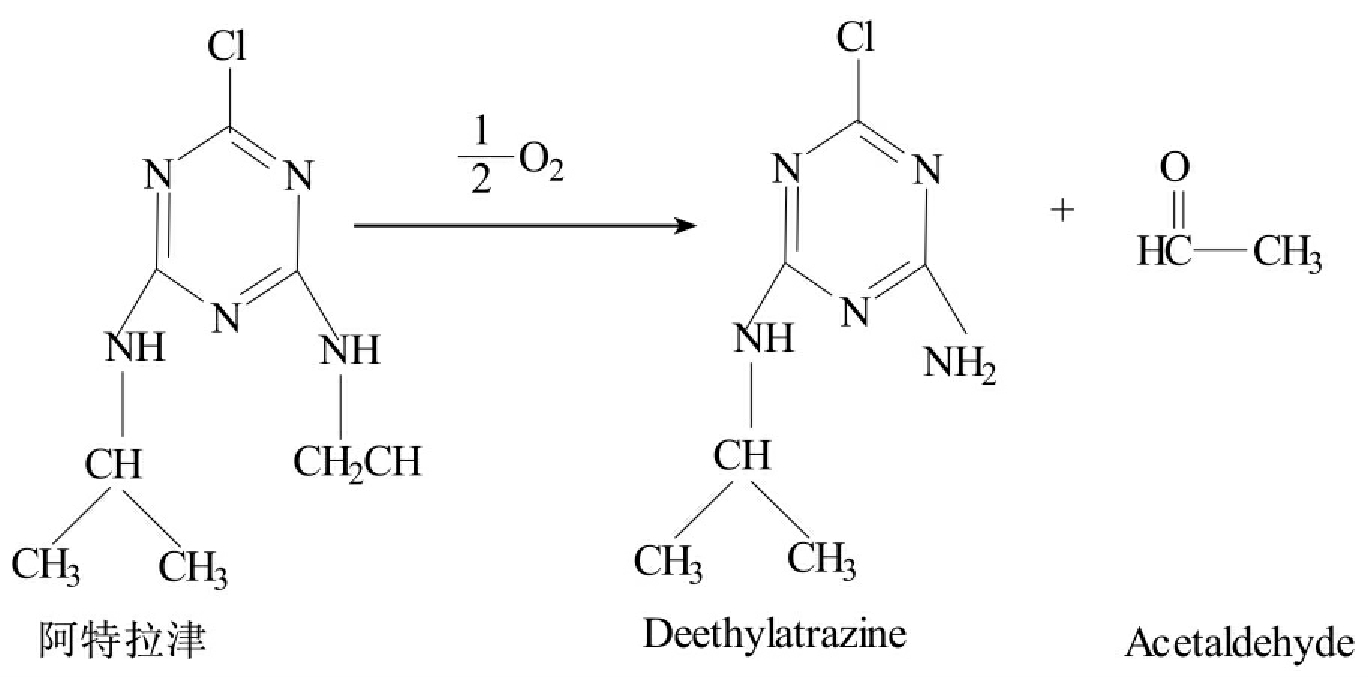
图8-11 阿特拉津的初始氧化

3.水解反应
水解反应是有机污染物降解过程中的重要反应,反应是向反应系统加水使底物中的某些基团发生水解。许多有机污染物的生物降解常开始于水解反应。有醚(ether)、酯(ester)或胺(amine)键的化合物在酯酶、丙烯酰氨酶、磷酸酶、水解酶和裂合酶催化下生成醚、酯、磷酸脂或胺键。此外还可以发生水解脱卤,卤素原子被水产生的羟基取代。这种反应也被称为取代反应,这如下式反应。
CH3CH2CH2Cl+H2─→O CH3CH2—CH3OH+HCl
重要农药阿特拉津降解的第一步也是水解脱氯反应,且其产物不具有原来农药的活性。
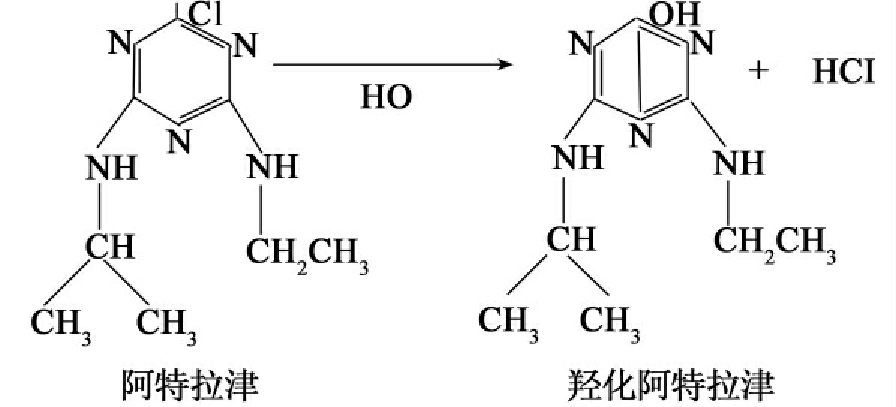
4.聚合反应
有机污染物通过增加某些化学基团扩增,形成聚合产物,或者把污染物接到另一个分子或多个分子得二聚或多聚化合物。在污染物的微生物降解中,常发生甲基化(methylated)、乙酰化(acetylated)和甲酰化(formylated),聚合以后的产物毒性比亲本化合物要小,然而在某些情况下,聚合后的污染物实际上对人和高等生物的毒性更大。
部分或者不完全降解也能导致聚合或合成比母体化合物更复杂、更稳定的化合物。这种情况发生在最初降解步骤,常常是胞外酶催化,并产生活泼的中间化合物,然后这些高度活泼的中间化合物能被结合或与环境中的有机物结合。例如除草剂敌稗(propanil)的水解产物3,4-二氯苯胺(3,4-Dichloroaniline)能被二聚化成为3,4-3'4'-四氯苯醌,三聚化成为4-(3,4-二氯苯胺)-3,3',4'-三氯偶氮苯(4-(3,4-dichloroanilino)-3,3',4'-trichloroazobenzene)。
二、生物降解途径规律性
微生物对有机污染物的降解利用实际上是微生物对许多物质分解利用能力的扩展和延伸。从微生物对糖类物质的分解利用过程中我们可以看到清晰的降解途径。
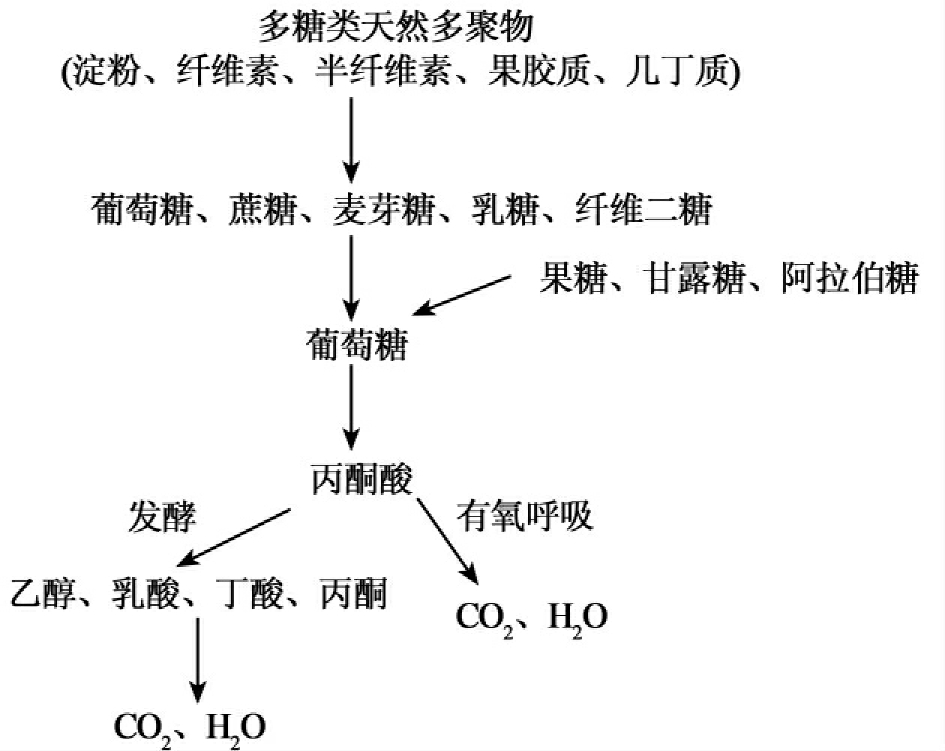
从中可以看到糖类物质分解途径的中心途径和旁支途径。中心途径是糖类物质从复杂多聚物产生的单糖,经葡萄糖、丙酮酸被进一步氧化成CO2和H2O。旁支途径是不同的微生物以及不同条件下所产生的代谢途径,在局部上所发生的明显改变。在污染物的生物降解中,芳香烃化合物是环境中最常见的污染物,主要来源于石油烃、溶剂、农药等方面。芳烃化合物因其结构的复杂程度不同经过不同的降解步骤后产生儿茶酚或取代儿茶酚,它们再经邻位裂解或对位裂解,而后产生丙酮酸,丙酮酸进入三羧酸循环而被彻底降解。它们的降解过程可以表示为:
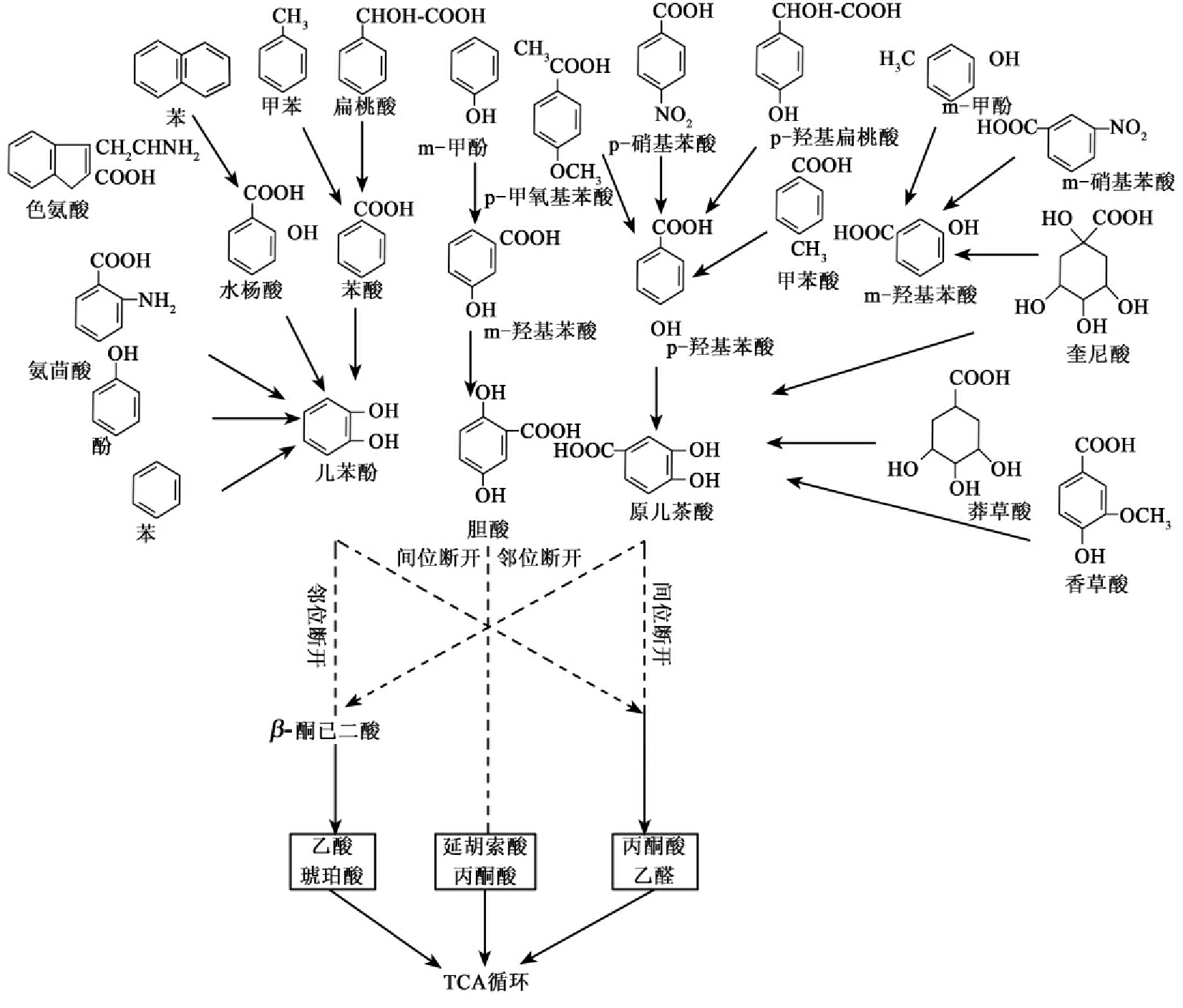
深入剖析芳香烃化合物的降解途径我们可以看到和前面所说的糖代谢降解途径有一致的模式,同样具有中心途径和旁支途径。儿茶酚、取代儿茶酚以后的降解途径可以认为是中心途径,而前面的可以理解为旁支途径。这是因为生物降解的整个进化实质上是对异生物源物质形成的新降解途径与已有的降解途径的接轨,如已在前面介绍的2,4-D的(2,4-二氯酚乙酸)的第一步降解产物2,4-二氯酚是自然界中存在的,已存在相关的降解途径,因此微生物对2,4-D的完全降解实际上只需进化出降解途径中第一步的催化酶(2,4-双加氧酶)即可。因此,环境污染物降解途径形拟为由无数支流汇成的一条大河。中心途径是主流,而旁支途径是支流,可表示为一种河流状结构。
三、降解速率及其动力学模型
1.降解速率
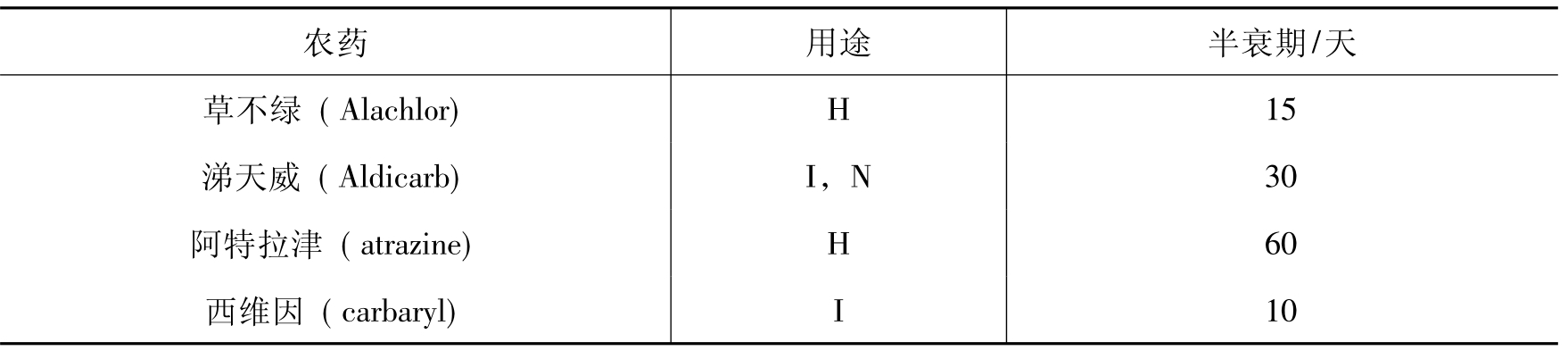
有些环境污染物是非常持久的,而有些则不是。降解速率是决定污染物在环境中归宿的关键因素。表8-5列出了17种广泛使用农药的平均半衰期值,其范围从5~2000天。DDT是高抗性化合物,半衰期达2000天。而甲基对硫磷(methyl parathion)和EPTC是相应非持久的,其半衰期分别为5~6天。
有机污染物的降解速率(degradation rate)有三种不同的表示方式,包括矿化速率(mineralization rate)、母体化合物消失速率(parent chemical disappearance rate)或总有毒残基(TTR)消失速率(total toxic residae(TTR)disappearance rate)。矿化速率代表一种有机污染物被完全降解成CO2、H2O和无机离子,一般污染物降解时测定放出的CO2代表矿化速率。为了区分环境介质中有机物呼吸放出的CO2,实际测定的污染物用放射性14C标记,这样放出的14CO2就代表了污染物的矿化速率,这样可以避免环境介质有机物降解的放出的CO2的干扰。母体化合物消失速率代表污染物从环境的消失。然而有些污染物的代谢产物有与母体化合物同样的生化活性(或生物毒性),并存在于环境中,在这种情况下,最好是测定TTR消失速率,测定出来是包括和母体化合物一样的有毒代谢物,而不是简单的母体化合物。
在有机污染物的降解过程中,并不是所有的有机碳都被矿化成可放出的CO2。部分有机碳将会掺入到微生物量,某些将会以有机代谢物形式积累下来,某些会转化成和土壤颗粒的结合残留物(bound residues)。这样有机污染物的矿化速率低于母体化合物的消失速率。由于TTR消失包含有毒代谢物,因此其速率将低于母体化合物的消失速率,但高于矿化速率。表8-6列出了三种农药的三种速率的半衰期值。Fenamiphos和涕灭威可以产生更毒的代谢产物,结果Fenamiphos和涕灭威的TTR半衰期值比相应的母体化合物消失值大得多。而虫螨威的两种值非常接近。
2.降解动力学模型
有机污染物在环境介质中的降解过程十分复杂,可以用不同的模型来表达。最好的方法是对所实测的数据进行拟合,而后得到最切合实际的动力学模型。
指数速率模型是最常用的用于描述降解过程的动力学模型,表达式如下:
-dc/dt=K1c
c为污染物的浓度(mg/g基质),t是时间(天),
K1为一级速率常数(1/天)
这种模型也被称为一级反应动力学模型。
根据这种模型,一种污染物在环境介质中的半衰期能用下列公式测定:
t1/2=0.693/K1
这里t1/2是污染物的半衰期(天),上面(表8-5)列出的半衰期值是在假设一级反应的动力学基础上得到的。
另一种常用的模型是双曲线速率模型,表达式为:
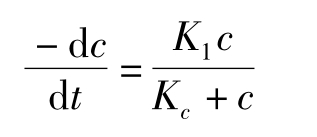
c为污染物浓度,K1为最大反应速率(mg/(g·天)),而KC则为最大反应速率一半时的浓度(mg/g)。
表8-5 某些常用的农药在土壤中的平均半衰期
续表
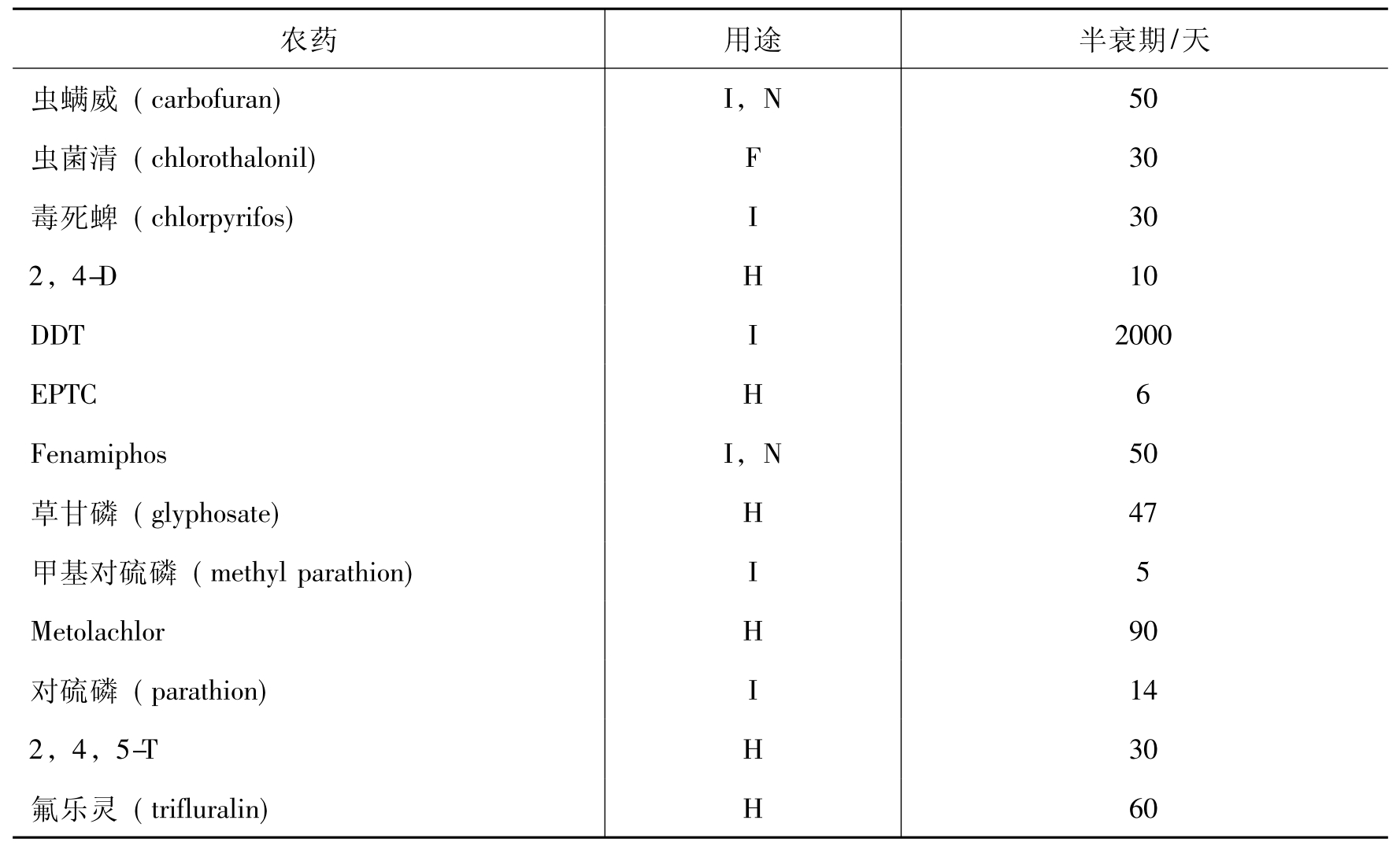
F:杀真菌剂H:除草剂I:杀虫剂N:杀线虫剂
表8-6 按三种降解速率计算的几种农药的半衰期

四、环境有机污染物在环境中行为
环境污染物在环境中的行为就是它们在生物降解过程中的去向。概括起来主要有三个方向。①环境污染物的彻底氧化,降解产物以气体或其他形态重新进入环境,如降解的终产物CO2、N2进入大气,以H2O形式溶入环境。②降解过程中的中间代谢产物被综合进入细胞,成为微生物生物量。③未被完全降解而残留在生态环境中。
1.环境污染物的彻底氧化
还原态化合物的彻底氧化及氧化态化合物的再合成是推动元素生物地球化学循环的两个重要过程。环境污染物完成降解产生的CO2、HO2以NO3-、PO4-可被光合生物进一步利用进行光合作用,合成有机物。环境污染物的降解过程推动了化合物的循环。
2.中间代谢产物被综合成微生物生物量
微生物在利用营养物过程中,产生的能量和中间代谢产物用于合成新的大分子物质(如蛋白质、核酸等),形成新的微生物生物量。一般情况下,微生物利用易于分解有机物时的生长是一种平衡生长,即底物有机碳量各有一半被氧化成CO2和被掺入细胞生物量。但环境污染物作一种底物被微生物降解利用时掺入生物量的有机碳的比例远少于对一般有机物的利用。在处理污水和固体废弃物的过程中,我们一般希望以较少的生物量处理尽可能多的环境污染物,因此在污水处理中减少污泥量,提高污泥的生理活性仍然是一个重要的研究课题。
3.环境污染物的残留
环境污染物在环境中的降解受到诸多因素及环境条件的影响,同时在与其他易于利用基质共存时,它们一般被滞后利用,因此实际上的完全降解在短期内很难达到。14C标记农药研究表明,仅有1%~50%的14C活性被放出。很多污染物的降解产物可与环境介质相结合,这就是污染物的残留。残留主要有结合残留。这些结合物不能用极性和非极性溶剂抽提出来,因此这些结合物也称为非可抽提14C活性。研究表明许多环境污染物(特别是农药)降解产物酚类化合物、氨基化合物或anilines可以共价结合到腐殖酸、棕黄酸、腐殖质上,这些称为结合残留物(bound residues)。在土壤中的结合残留物相当稳定,很难被降解。同时这种结合残留物一般可以大大降低其生物活性。对农药来说这种农药残留物对靶和非靶生物是无害的。另一种残留称为老化残留(aged rdsidues)。进入环境的污染物和环境介质相互作用,这种相互作用在土壤环境中的过程尤其复杂。在土壤中的相互作用包括土壤表面的吸附,溶解到土壤溶液,挥发到土壤孔隙空间,通过大孔和微孔扩散到内部土壤基质中,和仍然作为固体留在固体表面。可被微生物降解的部分主要是液相的部分,被吸附的部分一般难以被降解,当它们进入液相又可被降解,这又成为生物可利用的。对于那些疏水性农药,降解速率依赖于从土壤解吸的速率和程度,以及取决于从固相或气相溶解到液相。农药扩散到土壤基质内部,特别是扩散到微孔中,这种扩散主要依赖于化合物的挥发性和溶解性。这个过程被称为“老化”(aging)。这些残留下来的污染物(主要为农药)难以被微生物攻击,不被降解,因此这部分残留常被称为“老化残留”(aged residues)。老化农药残留物不能被有机溶剂在常温下抽提出来,但提高温度时可以抽提出来。老化农药残留物可以构成最终农药对地下水污染。
五、降解与解毒作用
许多环境污染物对人类及高等生物具有强烈的毒性,生物降解可以用于去除有毒化合物的毒性。微生物转化有毒化合物成为无机自然分子,如CO2、水、甲烷、硝酸盐和硫酸盐,结果最终使污染物脱毒,生物降解是脱除有机物毒性的有效方法。脱毒作用也可以发生在没有完全矿化的情况下。但有时降解过程中有些产物可能事实上具有比开始的分子有更强的毒性,或可以持久存在和积累在生境中。例如在厌氧条件下,四氯乙烯被代谢产生氯乙烯(vinyl chloride),这种产物不仅毒性非常高,而且也抵抗进一步的生物转化。这样对这些不完全降解的代谢产物我们更关心的应是证明其是无毒的。然而也应注意到即使最终被矿化的污染物也可能被转化成有毒的中间产物,这些中间产物也可以短时积累到可能的毒性水平。
许多毒性很强的有机物可以强烈抑制微生物的降解作用,但通过加入一些具有很强的吸附能力的多聚物(如环糊精)可以降低其生物毒性并促进生物降解,在有毒有机物的污水处理中这种方法可被利用。
六、环境污染物的酶促降解
对进入环境的合成化合物的转化取决于微生物酶对这些非生物源化合物的认识和对稳定性结构(如卤代有机物的碳卤键)的催化反应。环境污染物的抗降解性(顽固性)部分是由于酶的活性和酶专一性的问题。降解酶对环境污染物的降解是十分重要的。卤代脂肪族化合物结构多样,应用广泛,如用作起泡剂(氯甲烷)、冷却液(氯乙烯)、土壤薰蒸剂(1,3-二氯丙烯、溴甲烷)、杀虫剂(六氯环己胺)。化学合成中的中间产物(1,2-二氯乙烷、氯乙烯、氯乙酸)和溶剂(三氯乙烷、三-和四-氯乙烯)。各种氯代烷烃化合物也见于生产过程的废弃物中。脱卤酶受到广泛而深入的研究,这里以脱卤酶为例来说明降解酶的专一性、反应动力学及酶的工程化改造。
1.脱卤酶酶催化脱卤
有学者把酶催化脱卤的方式归纳为氧化脱卤、脱卤化氢、取代脱卤、甲基转移脱卤和还原脱卤等五种。
(1)氧化脱卤(oxidative dehalogenation)
氧化脱卤在卤代脂(肪)烃和卤代芳香烃化合物的降解中十分重要,其脱卤过程是由单加氧酶和双加氧酶完成的,而其中有的反应是共代谢完成的,而有的反应则是以正常的代谢方式完成的。卤代烯烃、卤代短链烯烃和某些卤代芳香烃化合物可以通过共代谢方式实现酶催化降解(参阅后面的共代谢部分及图8-15),具有这种功能的是单加氧酶和双加氧酶。目前已知的有甲烷单加氧酶、氨单加氧酶,酚单加氧酶,2,4-二氯酚单加氧酶、甲苯单加氧酶、甲苯2,3-双加氧酶、丙烷单加氧酶、烯烃单加氧酶和异丙基苯双加氧酶。除了共代谢外许多微生物产生的单加氧酶、双加氧酶也能直接氧化卤代化合物脱卤。如假单胞菌CBS3菌株产生的4-氯苯乙酸,3,4-双加氧酶能氧进行氧化脱卤(图8-12A)。
(2)脱卤化氢
脱卤化氢酶从卤代有机底物消除HCl,导致形成双键。少动鞘氨醇单胞菌(Sphingomonas paucimobilis)UT26矿化农药γ-六氯环己烷(六六六)时会发生脱卤化氢(图8-12B)。
(3)取代脱卤(substitutive dehalogenation)
取代脱卤包括水解脱卤、硫解脱卤和分子内取代脱卤三种。
①水解脱卤(hydrolytic dehalogenation):被研究的脱卤酶大部分是水解脱卤酶。卤代的杂环类、芳香类和脂(肪)环化合物的水解脱氯已有许多研究报道。最先被分离纯化的卤代脂(肪)烃脱卤酶来源于自养黄色杆菌菌株GJ10。这种细菌可以利用1,2-二氯乙烷作为碳源(图8-12C)。
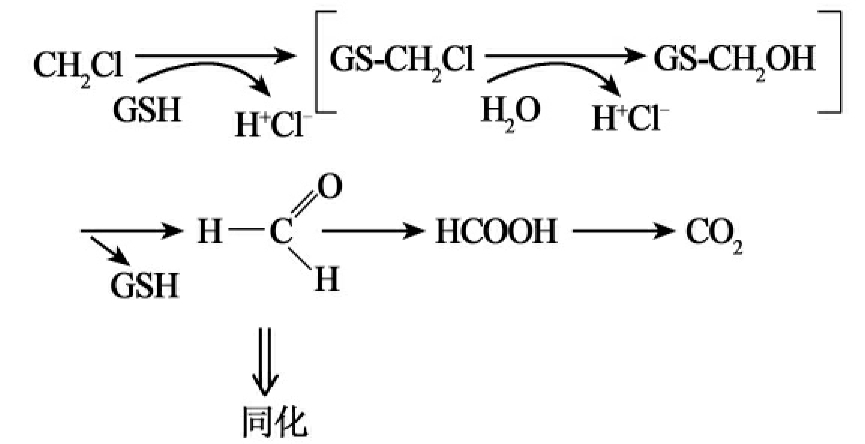
②硫解脱卤(thiolytic dehalogenation):谷胱甘肽S-转移酶催化形成氯化物和S-氯甲基谷胱甘肽。这种不稳定的中间产物被水解成谷胱甘肽,氯化物和甲醛,甲醛是甲烷营养菌生长的中心代谢物(图8-12D)。
③分子内取代脱氯(dehalogenation by intramolecular substitutive):卤代醇卤代氢裂合酶(Halohydrin hydrogen-halide lyases)(如卤代醇脱卤酶(haloalcohol dehalogenases)专一性催化邻位卤代醇和卤代酮的分子内取代反应,得到环氧化物。2,3-二氯-1-丙醇、1,3-二氯-2-丙醇、3-氯-1,2-丙二醇和其他卤代醇的降解(由假单胞菌菌株05-K-29、AD1等进行的)被认为是这种分子内的取代(图8-12E)。
(4)甲基转移脱氯(dehalogenation by methyl transfer)
甲基营养同型产乙酸(homoacetogen)脱卤醋酸杆菌(Acetobacteriamdehalogenans,即菌株MC)能利用氯甲烷作为唯一能源,产生乙酸。可被氯甲烷诱导的氯甲烷脱卤酶能转移作用底物甲基基团到四氢叶酸,产生甲基四氢叶酸和氯,甲基四氢叶酸被进一步代谢成乙酸盐,并通过乙酰-COA途径进一步反应(图8-12F)。
(5)还原脱氯(reductive dehalogenation)
还原脱氯对高氯代化合物(如PCDD和PCDF)的降解是十分关键的反应过程。
①共代谢还原脱氯(co-metabolic reductive dehalogenation)
许多产甲烷、产乙酸、硫酸盐还原和铁还原细菌能进行共代谢脱氯。这种共代谢主要见于卤代脂(肪)烃。反硝化假单胞菌菌株KC及腐败希瓦氏菌(Snewanella putrefaciens2000)菌株2000能通过共代谢降解三氯甲烷。这个过程受控于呼吸电子传递链的电子载体。细胞色素C、维生素K12被认为参与三氯甲烷的还原。
②耦联碳代谢还原脱氯(reductive dehalogenation linked to carbon metabolism):还原脱氯反应并不严格限于的厌氧细菌,紫色非硫细菌的深红红螺菌(R.rubrum)、度光红螺菌(R.photometrium)和血色红假单胞菌(Rhodospseudomonas rutila)在有氧条件下利用C2和C3卤代羧酸光能生长时,卤素取代基的还原是伴随着相应的羧酸的同化而进行的。
③呼吸过程中的还原脱卤(reductive dehalogenation as a respiratory process):许多菌株能把还原脱卤和能量代谢耦合在一起,它们以卤代脂(肪)烃或卤代芳香烃作为电子受体还原脱氯。一种硫还原细菌蒂氏脱硫念珠菌(Desulfomonile tiedjei)利用甲酸盐或H2作为电子供体,3-氯苯(甲)酸盐作为末端电子受体还原脱氯。还原脱氯和ATP合成的化学渗透假设的耦联(chemiosmotic coupling)已被证明。一种命名为Dehalococcides ethenogenes strain 195的细菌利用H2作为电子供体和PCE(五氯乙烷)为电子受体,完全脱氯PCE成为乙烯。PCE和TCE还原脱氯酶都是膜结合酶,并含有类咕啉辅因子。
A.
B.
C.
D.
E.
F.
图8-12 部分脱卤酶的脱卤反应
氯代烷烃脱卤酶催化C-卤键水解切开(cleavage)(图8-11C)的最先例子是自养黄色杆菌(Xanthobacter autotrophicus)和水生弯杆菌(Ancylobacter aquaticus)对1,2-二氯乙烷的降解,降解的第一步反应是由35kU的脱卤酶催化的,催化产生2-氯乙醇和氯化物。这种酶的底物范围很广,还能降解1,2-二溴乙烷,1,3-二氯丙烯以及长链氯烷烃等很多环境污染物。
卤代烷烃的降解酶中具有水解功能的是一组酶,一般属于α/β-水解酶,它们的活性中心中都有一个具有亲核攻击功能的ASP(天冬氨酸)残基。
自养黄色杆菌的脱卤酶(dehalogenase DhlA)的三维结构已被用x-射线晶体衍射法进行了分析。酶的活性中心是一个疏水的穴,位于具有α/β-水解酶折叠结构的球状主域(globular main domain)和分离的帽状区域之间。用x-射线晶体衍射法和同位素掺入法研究了催化原理。实验结果说明DhlA催化卤代化合物的断开是通过ASP124的羟化亲核取代(nucleophilic displacement)所进化的。产生的共价烷基-酶中间产物被活性水分子水解。水位于被脂化的ASP124的羰基碳上,HIS289以及ASP260通过减少一个质子使水分子活化。两种色氨酸包含在与底物卤素的结合中,面卤原子随后从底物中释放出来。
1,2-二氯乙烷是一种不存在于自然界的人工合成化合物,大量研究证明现在降解化合物的酶是近50年从一种更加原始的脱卤酶进化而来。支持其是最近进化而来的证据包括:①编码酶的基因的序列具有序列重复。②脱卤酶基因的组成型表达(黄色杆菌属和弯杆菌属菌株)。③基因定位质粒和从不同生物中分离到1,2-二氯乙烷脱卤酶缺乏进化趋异性。
2.酶的专一性
任何酶都有其底物专一性,酶的专一性分为两种类型,包括结构专一性和立体异构专一性。结构专一性按酶对底物专一程度的不同分为:绝对专一性(absolute specificity),酶对底物的要求非常严格,只作用于一种底物,不作用于任何其他物质。相对专一性(族专一性、基团专一性):酶作用底物不只是一种,对作用键两端的基团要求程度不同,对其中一端的要求严格,对另一个则要求不严格。键专一性酶只要求作用于一定的键,对键两端的基团并无严格的要求,这也是一种相对专一性,又称为“键专一性”,这类酶对底物结构的要求最低。立体异构专一性包括旋光异构专一性和几何异构专一性。旋光异构专一性是当底物具有旋光异构体时,酶只能作用于其中的一种。几何异构专一性是当底物具有顺、反两种结构时,只能催化其中一种结构,如延胡索酸水化酶只能催化延胡索酸即反-丁烯二酸水合成苹果酸,但不能催化顺-丁烯二酸的水合作用。
环境污染物的降解酶中有的是非专一性的,而有些有较强的专一性。共代谢转化是非专一性酶参与降解反应的重要例子,许多产生非专一性酶的微生物能降解非专一性的生理底物。如许多加氧酶不是专一性,如三氯乙烷可被发孢甲基弯菌(Methylosinus trichosporium) OB3b产生的溶解性甲烷单加氧酶(soluble methane monooxygenase)共代谢转化成一种环氧化物(epoxide),不稳定的环氧化物自发降解成各种产物。

而利用卤代脂肪烃作为惟一碳源和能源的微生物在利用性降解时的降解酶是专一性的。碳卤键的断开可能取决于酶的专一性,但单个酶的底物专一性不一定和化学结构直接相关。在以氯代脂肪烃化合物作为碳源方面,化合物的结构特征是重要的,从这个意义上这类化合物的降解酶从总体上说有专一性,并具有一定规律性(图8-13)。
3.酶催化反应动力学
氯代烷烃的脱氯酶的反应动力学受到较为深入的研究。研究者用稳态(steady state)和前稳态(pre-steady-state)相结合的动力学实验确立动力学原理和推算与此相关的速率常数(图8-14)。
使用中止一流荧光淬灭实验(stopped flow fluorescence quenching experiments)证明酰化的卤素离子从包埋活性中心穴中的释放制约整个酶的转化速率,而酶的构象改变可使卤素快速释放,构象改变是酶的帽状区域(cap domain)的部分运动,这种运动使水进入到活性中心,并使卤素离子溶剂化。
对脱卤酶转化1,2-二溴乙烷和1,2-二氯乙烷所有步骤的速率都作了测定,发现C-Cl键的打开速率要比C-Br键慢得多,这使二溴乙烷的Km值比二氯乙烷小得多。卤化物的释放是1,2-二氯乙烷和1,2-二溴乙烷水解速度的主要限制步骤(表8-7)。对许多其他底物转化的动态动力学分析说明,酶对溴化合物的亲和力明显高于氯化合物,这是因为对碳-Br键有较高的打开速率和对溴化合物有较高的二级结合速率常数(higher second-order association rate constant)。一般来说对于相应易于转化的底物在高浓度情况下,溴和氯化同系物以同样的速率转化,这是因为卤素释放所需要的构象改变是速度限制因素。而对于难转化的底物,溴化的同系物的转化要比氯化的同系物快得多,这是因为碳—卤键的断开是速率限制的。
4.酶的工程化改造
(1)卤代烷烃降解酶突变体的筛选
大量的研究证明许多降解酶的突变体可以提高卤化物的降解和卤素释放速率。脱卤酶Val226 Ala突变体对1,2-二溴乙烷的转化的Kcat值高出野生型1.8倍。但Kcat的增加并不见于对1,2-二氯乙烷的转化(表8-7),这可以解释为碳-卤键断开速率的明显降低,造成这种结果的原因是C—Cl键断开速率明显低于C—Br键,对突变体来说碳—卤断开成为对1,2-二氯乙烷转化的最慢的步骤。从这里也可以看到提高转化1,2-二氯乙烷酶的活性比提高1,2-二溴乙烷的酶活性要困难得多。提高活性的效应也见于卤代烷烃脱卤酶phe172Trp突变体(表8-8)。Phe172是螺旋-环-螺旋结构覆盖的活性中心穴疏水帽状区域残留基中的一个。Phe172Trp突变体酶比野生型对1-氯乙烷有10倍高的Kcat/Km值。研究表明其活性中心螺旋-环-螺旋结构的可塑性比野生型酶更高。这种增加使它更易结合1-氯己烷这样的大底物,有更强的亲和力,而野生型的活性中心穴则不适合于这种基质。对1,2-二溴乙烷转化速率的增加是由于在卤素释放前酶的异构化速率(rate of the enzyme isomerzation)的提高,这与前面提到的Val226Ala突变体情况一样,而异构化可能实际上是帽状区域部分构象的改变。
图8-13 结构特征和微生物可以利用作为生长基质的卤代化合物的关系破折线左边化合物能支持一种或多种微生物的生长,而右边则为难以利用的同系化合物。
图8-14 代烷烃脱卤酶的动力学模式
1.形成米氏复合物;2.碳卤键断开;3.共价中间产物断开;4.构象改变和卤素脱出
表8-7野生型卤代烷烃卤酶和Val226Ala突变体(226Val Ala突变体)四步反应的动力学常数
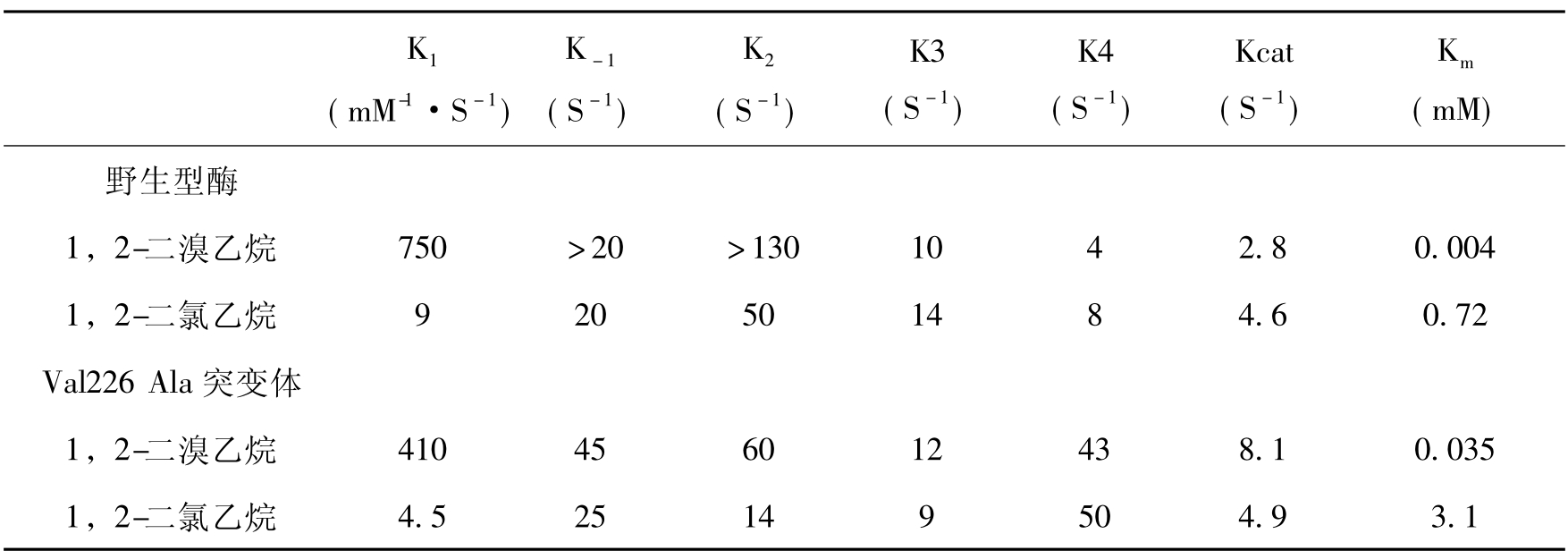
表8-8野生型、Phe172Trp(体外构建)和Asp170His(体内选择)卤代烷烃脱卤酶转化1,2-二氯乙烷、1,2-二溴乙烷和1-氯己烷的专一性常数(Kcat/Km值)

(2)新专一性的体内选择
通过对酶突变体的体内选择可以选择出降解原来不能降解底物的突变体,即为选择新的专一性,新专一性的体内选择。通过对自发突变的选择使脱卤酶适应新的底物,突变体能利用野生型酶难以水解的1-氯己烷。研究表明帽状区域的N端部分有短的定向重复,其对卤代烷烃脱卤酶适应新底物起重要作用。两个定向重复是15bp的优先重复和9bp重复。如果缺失两个重复就会失去1,2-二氯乙烷的水解活性,但仍具有对几种溴代脱卤酶底物的活性。这说明编码帽状的N端部分定向重复的DNA序列一定是最近进化出来的。而这个新的酶是在一个较老的脱卤酶是对工业产生的二氯乙烷适应中选择的。
(3)构建多酶体系的工程菌
对低分子卤代脂肪烃转化能力的瓶颈来源于断开碳-卤键和反应中间产物对生物的毒性。因此通过在合适生物中表达脱卤酶基因和构建带有多种能攻击顽固的结构和有毒的反应中间产物的酶的生物,可以提高微生物对化合物降解能力。有人已成功构建出基因工程菌,可以使极难降解的四氯化碳得到降解。
(4)利用已有的能转化非生物源化合物的有价值酶作为资源,开发具有区域和立体构型转化效应多样活性的酶。
(5)蛋白质工程的定位突变
定位突变技术是按照预先设计,精确地使靶基因在特定位点发生碱基序列的变化,进而使基因表达及其调控、基因产物发生相应改变。利用这种技术可以对目前的降解酶做进一步的工程改造,从而产生出具更强降解能力的酶。降解酶的定向改造和进化,见本章第一节。
七、降解菌及其降解基因的指示系统
把基因产物易于检测的报道基因(reporter genes)连锁到需要指示的降解基因的启动子上或整合到降解微生物的染色体(或质粒)上就可以构成降解基因的指示系统。利用这种指示系统,通过对基因表达的检出来评估降解基因及微生物的活性。这种系统依据的原理是大多数基因编码的产物及其活性不易检出,而一些可以作为报道基因的产物及活性却易于检出和定量测定。把报道基因连锁到降解基因或插入到降解微生物的载体上表达就可以检测到降解微生物的降解活性。报道基因的基本要求是易于检测,高度灵敏,可定量并能稳定表达(表8-9)。
表8-9 环境微生物中有用报道基因的特征

目前已经有多种基因被认定为可作为报道基因,主要包括代谢糖基因及发光基因,计有lacZ、gusA、xylE、luxL、lux(DABE)基因以及绿荧光蛋白(GFP)基因。
lacZ基因:基因编码β-半乳糖苷酶,它能把乳糖这种二糖切割成葡萄糖和半乳糖。在补加x-Gal的琼脂上生长的菌落产生蓝色,对其进行比色分析可定量分析β-半乳糖苷酶的切割产物,此外也可用荧光和化学发光试验来定量分析活性。但样品必须至少含有105~106个细胞才能定量分析。由于易于检出和有多种载体进行融合,lacZ是细菌培养物中基因调节研究的最常用的报道基因。
gusA基因:这种源于大肠杆菌的葡萄糖醛酸酶基因也是一种非常有用的标记,因为这种酶有许多荧光底物,而且从环境中分离出来的细菌很少能产生这种酶。许多底物被降解时能产生荧光,如5-溴-4氯-3-吲哚-β-D葡萄糖苷酸。gusA基因融合已广泛用于研究病毒性植物病原体、植物和真菌的基因表达,所有这些有机体缺少固有的β-D葡萄糖醛酸酶活性。这些条件使少到单个细胞的gus活性都能灵敏地测定。
xylE基因:xylE基因来自恶臭假单胞菌(Pseudomonas putida)的(降解)甲苯(TOL)质粒,编码儿茶酚2,3-双加氧酶。这种酶能把邻苯二酚转化成2-羟基黏康半醛,产生一种黄色色素,可用分光光度法测定。这个基因在没有接触过芳香烃污染物的微生物中很少见,使它有助于基因表达的系统的原位测试,已成功用于多种革兰氏阴性细菌。
发光报道基因在指示系统中的应用极为普遍,主要有lux和绿色荧光蛋白标记。
lux(lux CDABE)基因:这种基因编码荧光素酶及反应过程中的醛底物,luxAB编码活性荧光素酶,而luxCDE编码醛的合成酶。荧光素酶在作用底物荧光素时会发出荧光。把lux基因插入被研究的操纵子中,在那个操纵子被诱导时就会发光。这种基因在土生微生物中是极不常见的,因此已被广泛使用。大多数lux系统需要每个样品中含约105个细胞才能对发光进行检出,这种检出以一种不毁结构的方式直接进行,发出的光通常用闪烁计数器的单光子计数方式进行定量分析。这种基因在评估基因表达方面极为有用,并且已经用于评估异生物源有机物(如萘)生物降解。lux系统的应用还在不断扩展,光纤维埋进土壤可以用来指示微生物对土柱中污染物降解的原位反应。
GFP标记系统(绿色荧光蛋白标记系统):GFP基因见于维多利亚多管水母(Aequorea victoria)。GFP自身能把这种水母的蓝色生物发光转变成绿色,其原因还不清楚。当GFP基因在细胞(真核或原核的)中表达时,形成环状结构。紫外光(395nm)激发GFP会产生亮绿色荧光(509nm),这种亮绿色荧光可被测定,也可以作为细胞存在和数量的指标。与上述的lux系统相比,因为单个细胞中就有充足的GFP,所以可以测定单个细胞的基因转录而不是集群细胞的转录。同时GFP荧光不需要细胞代谢,因此并不活跃生长的细胞也能表达,这可能是许多环境中的实际情况。另外GFP和lux之间还有一个重要的差别,lux表达一停止发光就会停止,而GFP蛋白只要保持完整就会继续发光。这种lux是一个比较可靠的实时的活性指示基因。
第六节 环境有机污染物的共代谢降解
共代谢是环境有机污染物生物降解的重要方式。一种情况是共代谢的基质和加入物质在结构上相似,加入物质可以促进基质的降解,而另一种是易于被利用的基质可以促进难降解物质的降解。如缺陷短波单胞菌(Brevundimonas diminuta、Pseudomonas diminata)的一个菌株生长在易于利用碳源(如酵母提取物、肽或蛋白肽)上时能水解杀虫剂对硫磷成为P-硝基酚。
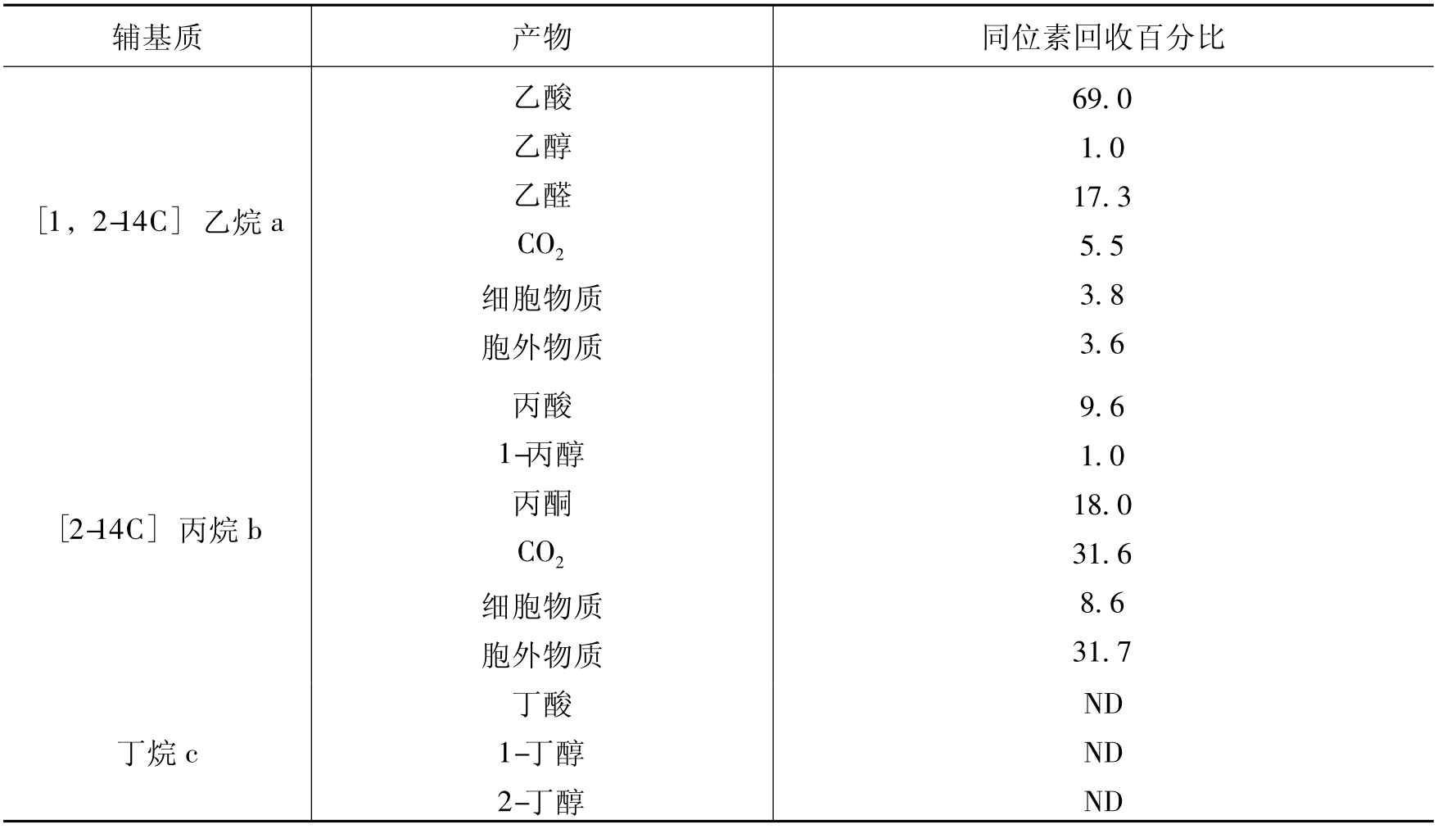
共代谢有多个意义相似,但又不完全一致的解释。①生长在一种可利用基质中的微生物可以同时转化(包括氧化)另一种并存但不能作为唯一碳源和能源的化合物。这种能使一种化合物转化(包括氧化)但不能成为碳源,取得能量和其他营养的现象称为共代谢或协同氧化。②只有在初级能源物质存在时,才能进行的有机化合物的生物降解过程。③微生物在利用生长基质时,可同时转化并存的但不能作为唯一碳源和能源的化合物的现象。④微生物转化一种不能利用作为能源和生长的化合物的现象。这种现象源于微生物酶的广谱专一性(broad-specificity),这些酶能认识与其酶促作用底物在结构上有相似之处的化合物,但酶促作用的产物一般不能被这种微生物的酶促反应进一步代谢。从某种意义上说共代谢实际上是微生物降解能力的水平延伸。共代谢的概念源于共氧化,研究者发现微生物在利用一种营养物质时能同时利用结构上相似的基质,那时就把这种现象称为共氧化。例如甲烷假单胞菌能利用甲烷,不能利用乙烷、丙烷和丁烷。但如果把甲烷和乙烷、丙烷、丁烷混合在一起时,甲烷假单胞菌在利用甲烷时,乙烷、丙烷、丁烷也同时被氧化(表8-10)。
此后许多科学工作者认为共氧化从本质上说就是一种共代谢过程,而且共代谢的概念范围更广,从此共代谢的概念被普遍接受。
目前共代谢的概念倾向在两种情况下使用,如定义②所说的在存在初级能源物质时,能进行原来所不能进行的有机物的生物降解过程。从这种解释出发,把那些不能为微生物作为能源和碳源的有机物或者那些难以降解的有机物投到一个含有大量易于微生物利用的有机物混合降解系统中,这些难以利用和降解的有机物的生物降解会得到促进,降解速率会得到提高,从而有利于这类有机物的去除,这对处理含难降解有机物的污水有重要的意义。但最普遍使用的共代谢是产生于酶的广谱专一性的代谢反应。研究得最深入、最具代表性的共代谢例证来源于甲烷营养菌(Methanotrophs)和其产生生物溶解性甲烷加单氧酶(soluble methane monooxygenase)。这个过程如图8-15所示。
表8-10 甲烷假单胞菌利用甲烷时对气态烷烃的共氧化
注:a气相组成:乙烷5%,甲烷45%,空气50% b气相组成:丙烷30%,甲烷40%,空气30% c未测定ND:未测定
一、共代谢降解
许多难降解有机污染物是通过共代谢开始降解而完成降解全过程的。这类污染物包括:稠环芳烃、杂环化合物、氯代有机溶剂、氯代苯环类化合物以及农药等。能进行共代谢降解的微生物包括好氧微生物、厌氧微生物和兼性微生物等。
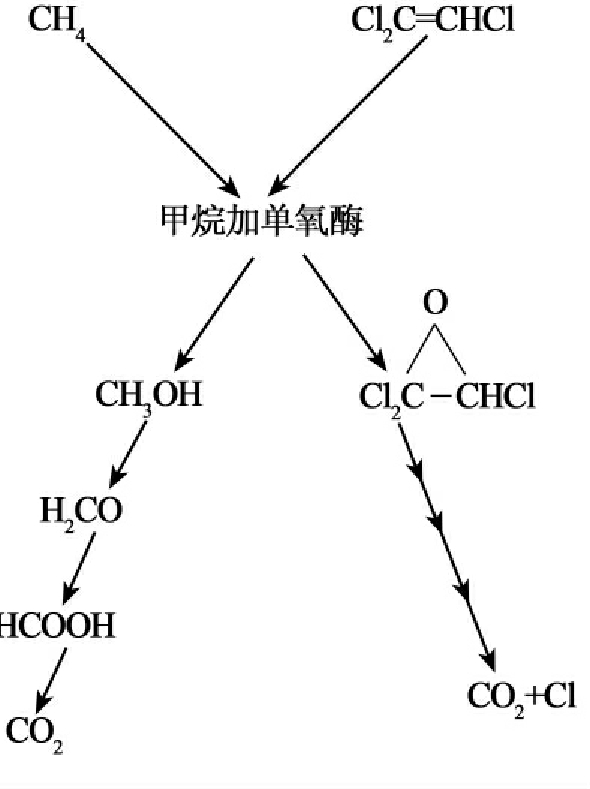
图8-15 甲烷加单氧酶催化的甲烷营养菌对TCE的氧化其后的降解步骤可能是由其他细菌或在某些情况下由甲烷营养菌自发催化的
三氯乙烯(TCE)的共代谢被深入研究,现以TCE为例简要说明共代谢的降解过程。TCE是一种很稳定的被广泛使用的工业溶剂,已经成地下水中最常见的环境污染物。TCE的分布广,其特别引人注目之处是TCE能被厌氧细菌还原脱氯成为氯乙烯(vinyl chloride),这种化合物对实验动物具有致突变和致癌性,也是人类的致癌剂。近百年许多科学工作者一直致力于得到能以TCE作为唯一碳源和能源的降解菌,但一直没有成功,由此TCE的共代谢成为人们关心的课题。至今已发现至少有9种细菌氧化酶能共代谢TCE,它们是:溶解性甲烷单加氧酶(Methylosinus trichosporiumOB3b)、甲苯2-单加氧酶(Pseudomonas cepacia G4)、甲苯4-单加氧酶(P.mendocina)、甲苯双加氧酶(P.putida)、氨单加氧酶(Nitrosomonas europaea)、颗粒性甲烷单加氧酶(Methylocystis parvus OBBP)、丙烷单加氧酶(Mylobacteriumsp.)、酚羟化酶(Alcaligenes eutrophus JMP134)、异戊二烯氧化酶(Rhodococcus erythropolis)。其中甲烷营养菌(Methylosinus trichosporiumOB3b、Methylococcus capsulatus)及其产生的溶解性甲烷单加氧酶,恶臭假单胞菌F1及其产生的甲苯加双氧酶受到深入的研究。溶解性甲烷加单氧酶在低铜浓度时合成,酶系由三种蛋白成分组成,羟化酶成分含有双核铁中心作为氧结合和反应位点。溶解性甲烷单加氧酶氧化TCE如图8-16所示。甲苯双加氧酶对TCE的氧化如图8-17所示。
甲烷营养菌氧化甲烷的第一步是由甲烷单加氧酶催化的,这种酶是广底物专一性的,其在甲烷和TCE同时存在的情况下也能共代谢催化TCE氧化过程的第一步。这种细菌并没有从共代谢步骤中获得能量上的好处。TCE共代谢的降解产物可以被其他细菌或许也可能是甲烷营养菌进一步催化降解。在修复被TCE污染的环境中,我们可以向环境中投入甲烷、甲苯、丙烷甚至氨这样的共基质(co-substrate),利用甲烷营养菌、甲苯利用菌、丙烷利用菌及氨氧化菌的共代谢能力来净化TCE污染环境。在纯培养条件下的共代谢是一种截止式转化(dead-end transformation)。然而在混合培养和自然环境条件下,开始的共代谢可以为其他微生物所进行的共代谢或其他降解铺平道路。以这种共代谢方式,使难分解的污染物经过一系列微生物协同作用而得到彻底降解。同时这种偶然转化中的共代谢可能有害,能导致对进一步降解具有更大抗性和毒性化合物的产生。
共代谢过程除TCE这种方式外,还可以有其他多样的方式,例如能利用DCA(1,2-二氯乙烷)作为唯一碳源和能源生长的假单胞菌DCA1能共氧化利用DCP(1,2-二氯丙烷)。用DCA作为辅基质共代谢DCP时会出现一种竞争性抑制,造成共代谢过程中降解微生物生长慢,降解速率低的问题。有研究表明DCA降解过程中的中间代谢产物乙酸易于被DCA利用菌氧化利用,加入氯乙酸能使细胞表达DCA单加氧酶,而又不与DCP竞争这种单加氧酶,使DCA单加氧酶可以共代谢DCP。
在氯代芳烃中,氧化脱氯对高氯化合物是无能为力的,如四氯乙烯就不能氧化脱氯,还原脱氯可以使高氯化合物实现脱氯,同样在还原脱氯中也存在共代谢,已经发现在甲烷产生菌的纯培养中氯乙烯和氯乙烷也发生还原脱氯。
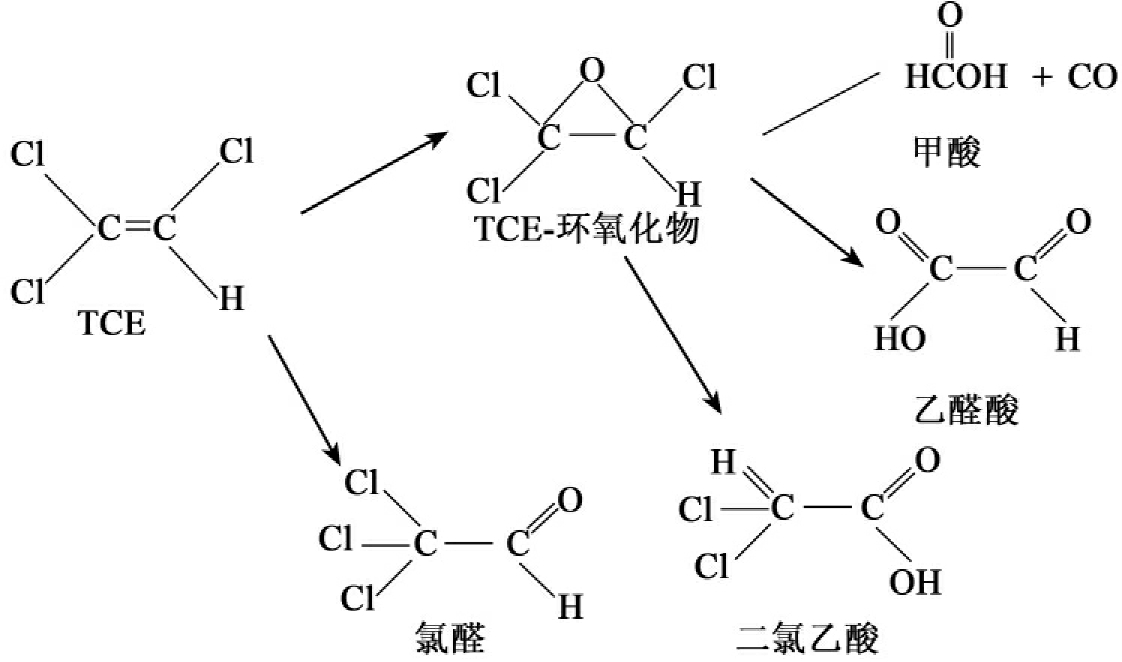
图8-16 溶解性甲烷单加氧酶对TCE的氧化

图8-17 甲苯加双氧酶对TCE的氧化
二、共代谢机理
许多难降解环境污染物的生物降解开始于氧化酶的氧化,而许多氧化酶具有广基质谱(broad subserate spectrum)的特征。这种特点使这些酶在氧化其专一性底物的同时也作用于化学上与其专一性底物相似的化合物。专一性底物在这个共代谢系统中被称为共基质(cosubstrate)和主要生长基质,被共代谢的底物称为共代谢底物(即难降解有机物)。能进行共代谢的酶被称为关键酶。共代谢降解的产物一般不能被产生共代谢降解酶的微生物进一步降解和利用。但可被其他的微生物降解和利用。关键酶可被辅基质或其他结构类似物所诱导。
三、共代谢降解的特点
共代谢降解过程的主要特点可以概括为:
1)微生物利用一种易于降解利用的基质作为碳源和能源,促进微生物的生长和产生数量较多的微生物物量。
2)有机污染物作为第二基质被微生物降解。有转化能力的微生物在只存在这种化合物时不能繁殖,不能生长,污染物的转化速率在整个时间不能增加,这样事实上,由于开始转化的种群小,降解能力比较低。
3)污染物与营养基质之间竞争降解酶的活性中心,浓度及亲和力是决定反应的关键因素。
4)污染物共代谢的产物不能作为营养被同化成为细胞质,有些对细胞有毒害作用。转化反应的产物可以积累在环境中。
5)共代谢是需能反应,能量来自营养基质的产能代谢。
6)共代谢反应由种类有限的几种活性酶——关键酶决定的。不同类型微生物所含有的关键酶的功能是类似的,例如,好氧微生物中的关键酶主要是加单氧酶和加双氧酶。
7)围绕关键酶的共代谢存在着极为复杂的调控关系。
①关键酶控制着整个反应节奏,其浓度由共基质(第一基质)诱导决定。共基质的诱导作用决定着共代谢中微生物体内关键酶的浓度,但由于共基质和共代谢底物之间存在一种对酶的竞争作用,高浓度营养基质反而导致降解速率的下降;能量基质能提供高降解速率,但是高浓度能量基质不利于长期维持微生物的降解活性(高能量导致共基质的急剧消耗,降低诱导酶的能力)。事实上共氧化氯代化合物对在混合培养中的细菌的竞争是一个劣势。共代谢氧化需要还原等价物,因此降低了用于生长的可利用能量。而且可能形成的有毒中间产物有损于生长。
②毒性中间产物抑制关键酶的活性。微生物能够迅速启动自我修复功能以对抗毒性抑制作用。
③微生物通过关键酶提供共代谢反应所需要的能量。
四、共代谢的应用
共代谢反应广泛存在于氯代有机溶剂、氯代苯环类化合物、卤代芳烃、杂环化合物以及农药等环境有机污染物的生物降解中,共代谢现象加深了我们对生物降解过程的认识,同时也在生物修复、净化环境中具有很大的应用价值。
①在难降解有机物生物降解试验中,不能单一设置以该物质为唯一碳源与能源的试验系统,应加入共代谢的共基质,采用污染物逐步消失试验。
②以共代谢原理指导人工合成材料的成分组成。如原来由聚氯乙烯组成的塑料是难降解的,现在国外研究加入淀粉物质使其易于为微生物降解。以淀粉作为能源,然后再对难降解物质作用。如美国研制的可自行销毁的塑料薄膜是由玉米淀粉(副产品)和聚乙烯构成的复合材料,在土壤微生物作用下可迅速被降解。又如把土豆屑(食品工业的重要废品)转化成乳酸(土豆屑→糖→乳酸)制成乳酸塑料也易于降解。
③在污水处理中通过添加初级基质来处理含难降解污染物的污水,使难降解物质得到生物降解。
④构建共代谢的生物降解系统修复难降解有机污染物污染环境。一般对不存在利用这种污染物作为惟一碳源和能源微生物情况下,修复过程可以向污染环境加入共基质诱导共代谢的关键酶代谢降解这种难降解污染物(具体应用见生物修复相关章节)。
⑤研究表明水杨酸、邻苯二甲酸等可以作为共代谢共基质加到石油烃污染环境中有助于修复污染环境。
⑥许多多环芳烃具有相似的结构,相互关系复杂,大量研究表明微生物对多环芳烃的降解中存在共代谢的关系,这样获得对一种多环芳烃降解能力的微生物就有可能共代谢结构相类似的一类多环芳烃的降解能力。
第七节 常见有机污染物的生物降解
目前有10万种以上的化合物被商业生产,其中有数百种被大量生产,这其中大部分是有机化合物,有机化合物的环境污染物给人类带来极大的风险,生物降解是降低,规避风险的重要途径。有机污染物种类多样,结构复杂,难以对它们作统一科学的分类。综观大量被公认的有机污染物,我们可以看到它们中的大部分结构基础是烃键和苯环,且最终的形态是从这个基础上衍生出来的,为了便于从结构上对它们的生物降解进行比较分析,并从中认识生物降解的规律,本章把常见有机污染物分类为烃类化合物和其他化合物。
现存微生物对有机污染物的降解能力是它们对结构类似的化合物降解能力的延伸和扩展,不同种类微生物的进化历程、生存环境以及对新有机物的接受驯化,以及获得新的降解能力途径不同,因此有机物的污染物的生物降解会因不同的生物种类,不同的微生物群体,不同的生态环境而不同,呈现出错综复杂的情况,可以有不同的降解途径。因此我们对有机污染物的生物降解的认识还是很初步的,这里介绍的只能代表其中的一种或数种。
一、烃类化合物的生物降解
1.脂肪烃化合物
进入环境成为污染物的脂肪烃化合物来源多样,包括石油烃中的直链烃、带支链烃、表面活性剂的烷基取代物,卤代一碳或二碳化合物(如TCE)。卤代物是普遍使用的工业溶剂。这类化合物的一般降解规律:①中等链长的直链脂肪烃(链长10到18个碳的直链烷烃)比更短或更长的更易于被利用。更短链长的烃有很高的水溶性,对细胞脂和膜具有损伤,破坏细胞的完整性。而链长更长的烃水溶性降低,因而降解性降低。②饱和脂肪烃和不饱和的烯烃降解性相当。③烃的支链会降低生物降解性。④卤素取代基会降低生物降解性。
(1)无取代基的脂肪烃化合物
①烷烃。烷烃与自然界普遍存在的脂肪酸、植物蜡结构相似,环境中许多微生物都能利用直链烷烃作为唯一的碳原和能源。大量能降解直链烃的微生物已从烃污染环境中分离出来,假单胞菌属、产碱菌属、芽孢杆菌属等许多菌都具有这种能力。好氧条件下直链烷烃生物降解的途径主要有四种。最常见的途径是加单氧酶把一个氧原子掺入烷烃末端的一个碳中生成伯醇,另一种情况是加双氧酶将两个氧原子掺入烷烃,生成过氧化物。此外也有双末端氧化和亚末端氧化,最终都生成脂肪酸,脂肪酸最后经β-氧化被彻底氧化分解。带有OCT质粒的食油假单胞菌(Pseudomonas oleovorans)降解庚烷的途径如图8-18。降解过程的酶是位于质粒上的alkBGTJKL编码的(alk源于alkane(链烃))。
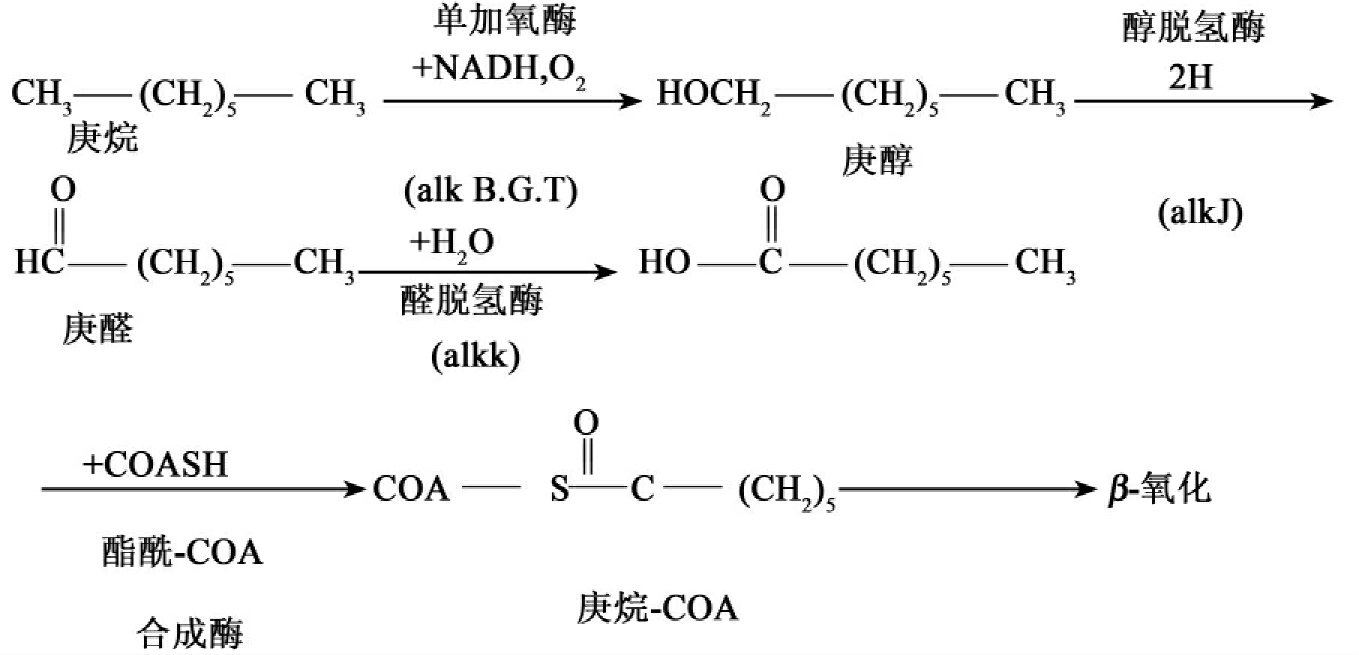
图8-18 携带OCT质粒的食油假单胞菌降解庚烷的途径
alKB(膜结合水解酶)、alKG(红氧(化)还原蛋白(rubredoxin))和alKT(红氧(化)还(原)蛋白还原酶)编码的多成分单加氧酶把烷氧化成醇。后两种蛋白构成一条短的电子传递链。alKJ和alKK催化醇的循序氧化产生醛和酸。alKL的基因产物把COA共价连接到酸性的羟基上得到庚烷COA,而后再经β-氧化被完全降解。
在严格厌氧条件下饱和脂肪烃降解缓慢,这一点有大量事实可以支持,在自然地下油库厌氧条件下尽管存在大量微生物,但不会被降解。
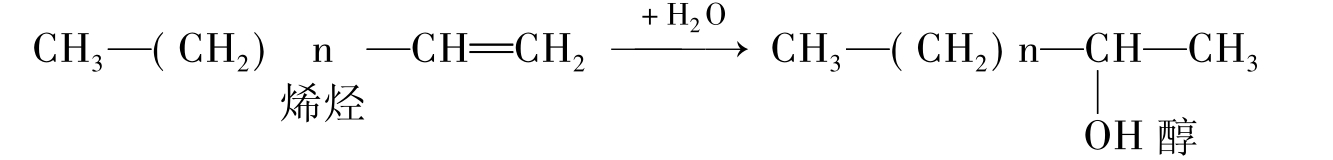
②烯烃。烯烃是在分子中含有一个或多个碳碳双键的烃。烯烃的生物降解速率与烷烃相当。图8-19以单烯为代表,好氧条件下的降解步骤包括对末端(1)或亚末端(2)甲基的氧化攻击,攻击方式如同烷烃。另外的开始步骤是攻击双键,产生伯醇(3)仲醇(4)和环氧化物(5)。而这些最初的降解产物都会被进一步氧化生成脂肪酸,并逐渐经β-氧化分解。在好氧条件下烯烃易于生物降解,降解途径是先将双键羟基化为醇,而后进一步被氧化为醛,最后转化为脂肪酸。
(2)卤代脂肪烃化合物
卤代脂肪烃化合物被广泛使用。像三氯乙烯(TCE)被广泛用作工业溶剂。由于使用和处置不当,这些溶剂是地下水中最频繁检测到的有机污染物之一,其生物降解受到广泛的研究。卤代脂肪烃的降解速度比没有卤代的脂肪烃慢得多,同一个碳原子键合二或三个氯原子则其好氧降解受到抑制,此外从C3到C12的一氯化烷烃的降解速率随碳链的加长而增加,这可以解释为随着链长的加长,氯原子对酶-碳反应中心的电子效应的减弱。
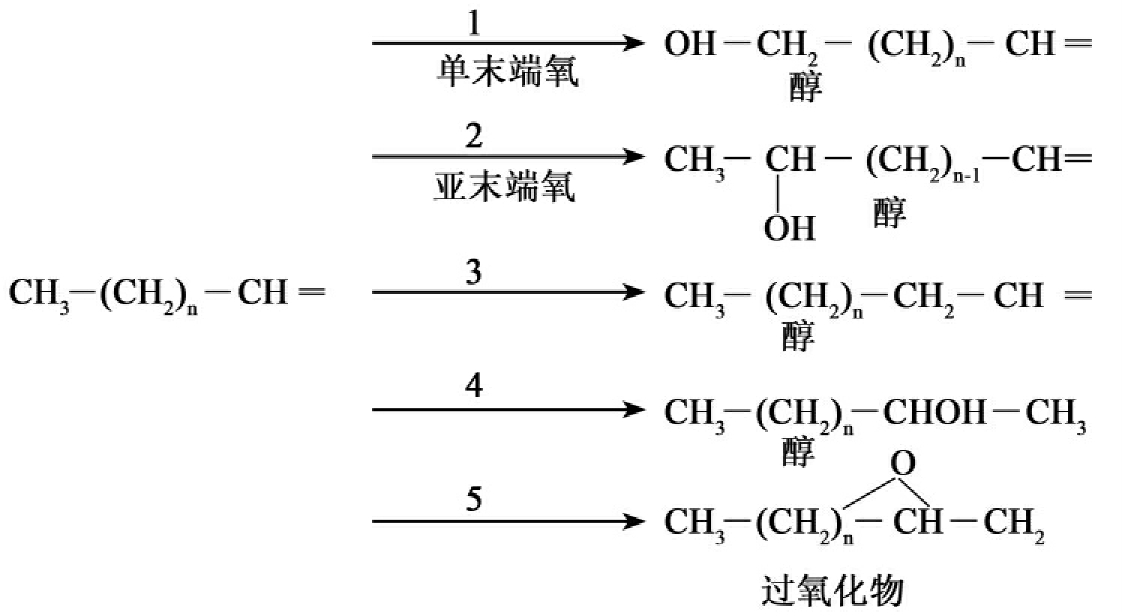
图8-19 烯烃的生物降解
卤代脂肪烃的好氧生物降解反应有两种基本类型:
①亲核的取代反应(反应物带来一对电子),一卤或二卤代化合物的卤原子被羟基取代(图8-20)。
②单加氧酶和双加氧酶催化的氧化反应,这些酶能氧化高度氯代的C1和C2化合物(如三氯乙烯)。能氧化包括甲烷、氨、甲苯和丙烷各种非氯代化合物的细菌可产生这些单加或双加氧酶。这些酶没有严格的底物专一性,它们能共代谢氯代脂肪烃(参阅本章第六节)。通常氯代脂肪烃的共代谢降解需要相对大比率的底物。研究证明以甲烷或甲酸盐为碳源生长的甲烷营养菌产生的甲烷单加氧酶,以甲苯为碳源的甲苯营养菌产生的甲苯双加氧酶,以氨为营养的欧州亚硝化单胞菌产生的氨单加氯酶和以丙烷为营养的母牛分枝杆菌SOB5产生的丙烷单加氧酶都具有共代谢降解卤代脂肪烃的能力。
卤代脂肪烃在厌氧条件下还原脱氯(图8-21)(参阅本章第五节的环境污染物的降解酶),电子由还原性金属转移到卤代脂肪烃上,产生一个烷基和一个游离卤原子。而烷基基团能吸引一个氢原子(1)或失去第一个卤原子形成一个烯烃(2)。
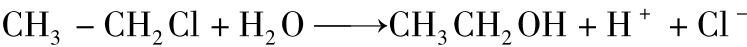
图8-20 卤代脂肪烃的取代脱氯
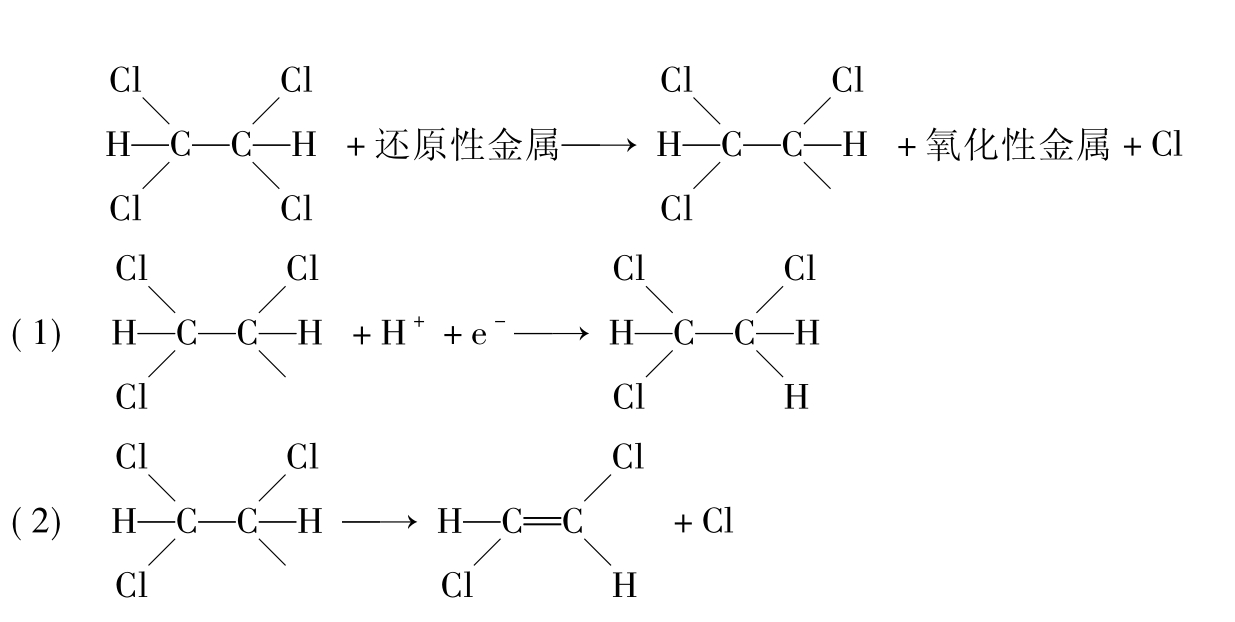
图8-21 四氯乙烷还原脱氯形成三氯乙烷(1)或二氯乙烯(2)
通常好氧条件有利于较少卤代取代基的化合物的生物降解,而厌氧条件上有利于较多卤代取代基的化合物的生物降解。然而在厌氧条件下,高度卤代的脂肪烃不能完全降解。因此开始厌氧条件下处理降低卤代程度,然后充氧创造好氧条件进行好氧过程使卤代物完全降解。
2.脂环烃化合物
脂环烃化合物在化学工业上的使用以及不包括开采和使用在内的工业过程所产生的对环境释放都是有限的。因此人类暴露在脂环烃化合物下的健康风险后果不像其他化合物(特别是芳香烃化合物)那样达到同样重要的水平。它们的生物降解研究注意得较少。
脂环烃没有末端甲基,它的生物降解原理和链烷烃亚末端氧化相似,以环己烷为例(图8-22)来说明。混合功能氧化酶氧化产生脂环(族)的醇,脱氢得酮。进一步氧化形成一个内酯。不稳定的内脂环断开得到羟基羧酸。羟基再被顺序氧化成醛基和羧基。得到二羧酸可被进一步氧化分解。有人从污泥中分离到一株诺卡氏菌能以环己烷作为唯一的碳源生长。然而更常见是这个降解过程是一个共生的共代谢反应。在这一系列反应中,一种微生物将环己烷经由环己醇转化为环己酮,但这不能使环己酮内酯化和开环,而另一种不能开始氧化环己烷的微生物却能使环己酮内酯化和开环,并进一步氧化分解。
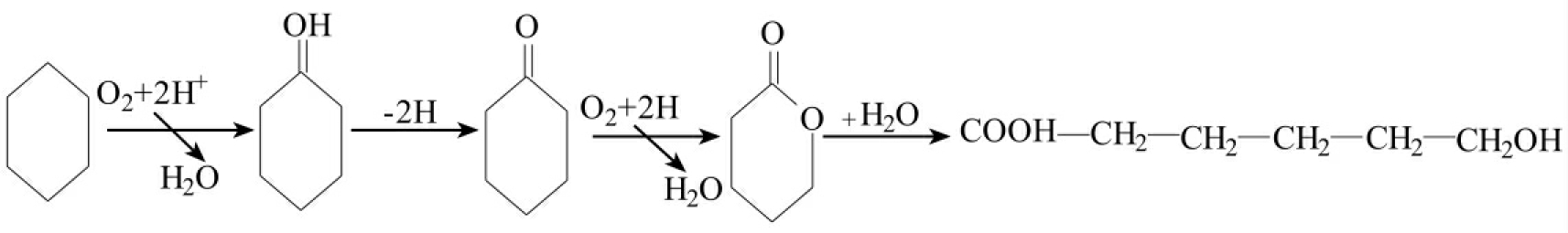
图8-22 环己烷的降解途径
3.芳香烃化合物
芳香烃化合物含有至少一个不饱和环状结构,通常是C6R6,R可以是任何基团。苯是这个不饱和环状化合物家族的母体烃,含有两处或更多合在一起的苯环化合物称为多环芳烃(PAH)。芳烃化合物的氢原子可以被许多基团所取代,从而形成取代基芳香烃化合物。芳香烃化合物既有自然来源的,但更多来自许多工业部门的废水、废弃物。由于对人类健康和生态系统的潜在毒性和影响,芳香烃化合物的生物降解被广泛研究。
(1)无取代基芳香烃化合物
研究表明大量的细菌和真菌能够在各种环境条件下部分或完全降解芳香烃化合物。在好氧条件下,最普遍的初始转化是把分子氧掺合到芳香烃的羟化作用。催化这种反应的是加单氧酶和加双氧酶。
苯是芳香烃的基本结构,多环芳烃降解最终也要经历到苯,并进一步转化,最终完全降解。苯的生物降解在芳香烃的生物降解中具有重要的代表性。
原核微生物对苯的生物降解如图8-23所示。在加双氧酶的催化下苯被转化为顺式二氢基二醇,然后这个二氢基二醇重新芳香构化形成一种二羟化中间产物儿茶酚。儿茶酚的环在第二个加双氧酶的作用下被打开,在两个羟基之间打开为邻位途径,在一个羟基的下一个位置打开为间位途径,此后可进一步反应直到完全降解。

图8-23 苯经儿茶酚的降解过程
真核微生物对苯的降解是用细胞色素P-450加单氧酶攻击芳香烃化合物,把分子氧的一个氧原子掺合到化合物中而另一个氧原子被还原成水,结果生成一种芳烃氧化物。接着在酶作用下与水加成生成反式的二氢基二醇。另一种情况下,芳烃氧化物能被异构化为苯酚,而苯酚能和硫酸盐、葡萄糖醛酸、谷胱甘肽缀合,这些缀合物能被排出(图8-24)。
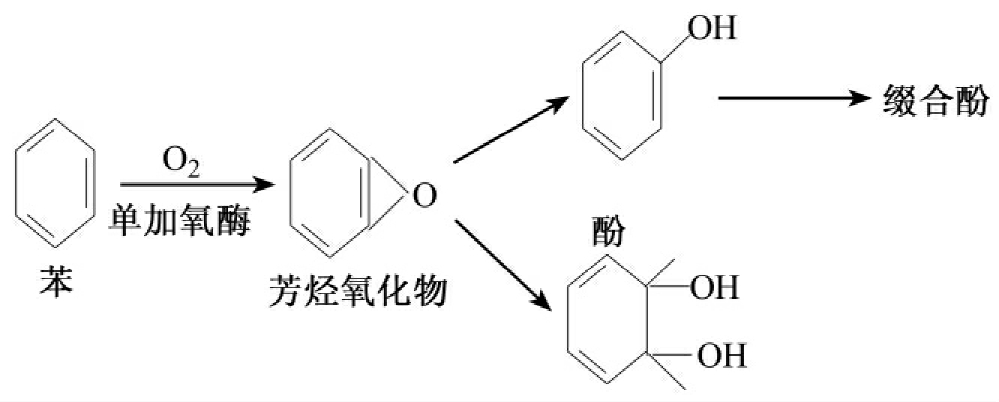
图8-24 真核微生物分解苯的过程
多环芳烃的生物降解过程十分复杂,一般来说二环(如萘)、三环的多环芳烃(如蒽、菲)研究得较为广泛深入,而更多环,更复杂的多环芳烃(如、chrysene和亚苄基芘(benzola pyrene))研究得相对较少,较不深入。总体上其降解是从攻击其一个环开始,而后再打开另一个环,最后成为单环化合物,单环化合物的生物降解类似苯的生物降解,最终完全降解。常见几种多环芳烃的酶攻击位点如图8-25所示。
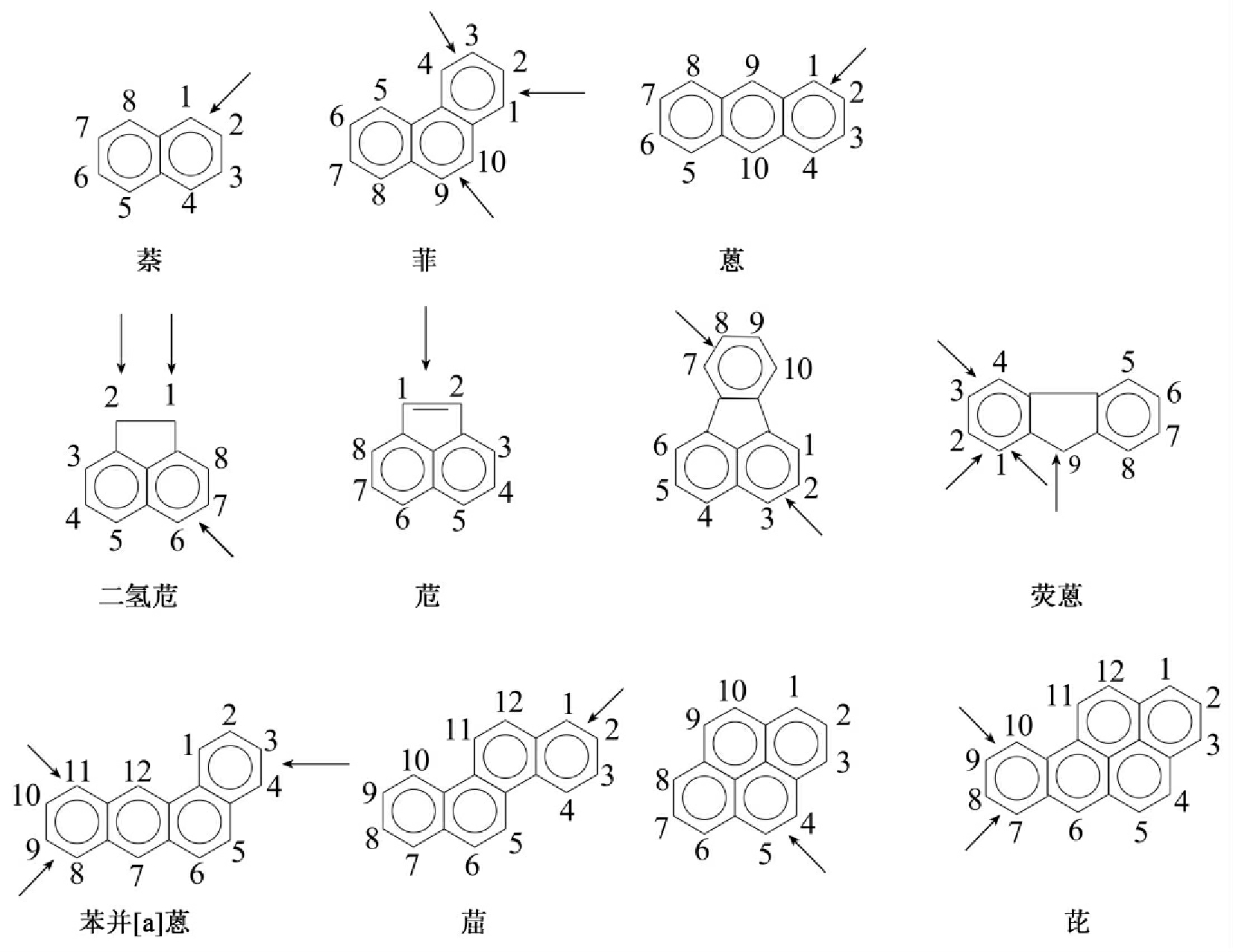
图8-25 多环芳烃酶攻击位点示意图
多环芳烃的好氧生物降解主要有三种途径,其一是被细菌和绿藻氧化成顺式二氢二醇(Cis-Dihydrodiols),再经苯酚、环断裂被降解。其二是被甲烷营养菌代谢形成苯酚。其三是被真菌、细菌和蓝细菌代谢形成反式-二氢二醇(trans-Dihydrodiols),然后进一步降解。
大部分细菌和某些绿藻在好氧条件下,可以通过加双氧酶氧化PAHs形成顺式二氢二醇,二氢二醇被进一步转化成二酚,再被其他的加氧酶打开,并进一步氧化降解。
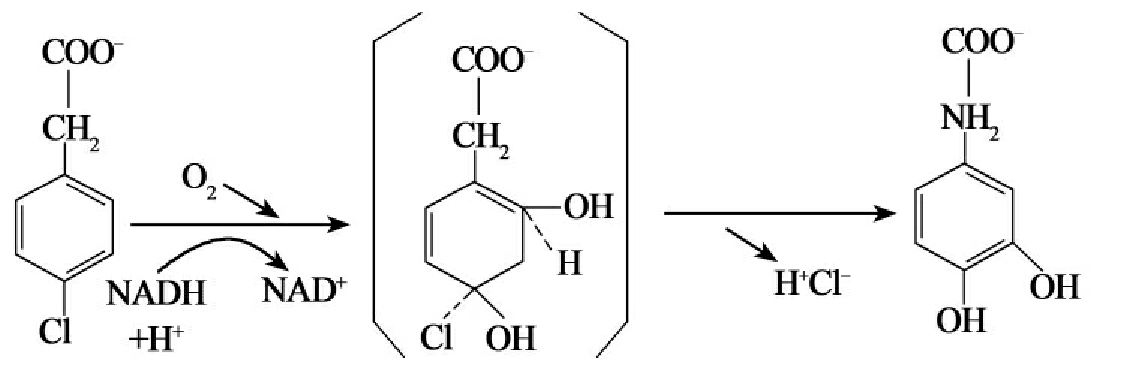
二环萘的降解及降解途径受到广泛研究。恶臭假单胞菌(Pseudomonas putida)降解萘的途径如图8-4所示。具有同样降解途径的还有乙酸钙不动杆菌(Acinetobacter calcoaceticus)及分枝杆菌属(Mycobacteriumsp.)、红球菌属(Rhodococcus sp.)的菌株。
铜绿假单胞菌(Pseudomonas aeruginosa)在好氧条件下降解三环蒽的途径如图8-26所示。这种细菌把蒽转化成蒽顺式-1,2-二氢二醇(cis-1,2-dihydrodiol)、1,2-二羟蒽(1,2-dihydroxyanthracene)、cis-4-(2-hydroxyo-aphth-3-y1)-2-oxobut-3-enoic acid、2-hydroxy-3-naphth-aldehyde和2-hydroxy-3-naphthoic acid。后者再被矿化成水杨酸和儿茶酚。荧光假单胞菌、红球菌的菌株也能利用蒽。
三环菲的生物降解途径与蒽相似,假单胞菌能代谢降解菲(图8-27)。菲被转化成菲顺式-3,4-二氢二醇(phenanthrene cis-3,4-dihydrodiol)、3,4-二羟菲(3,4-dihydroxyphenan-threne)、cis-4-(1-hydroxynaphth-2-y1)-2-oxobut-3-enoic acid、1-hydroxy-2-naphthaldehyde、1-hydroxy-2-naphthoic acid。后者再被矿化并通过萘的途径被降解。此外气单胞菌属、拜叶林氏菌属、分枝杆菌属和红球菌属、节杆菌属的一些菌株也能以相似的途径降解菲。

图8-26 铜绿假单胞菌等细菌代谢三环蒽的途径
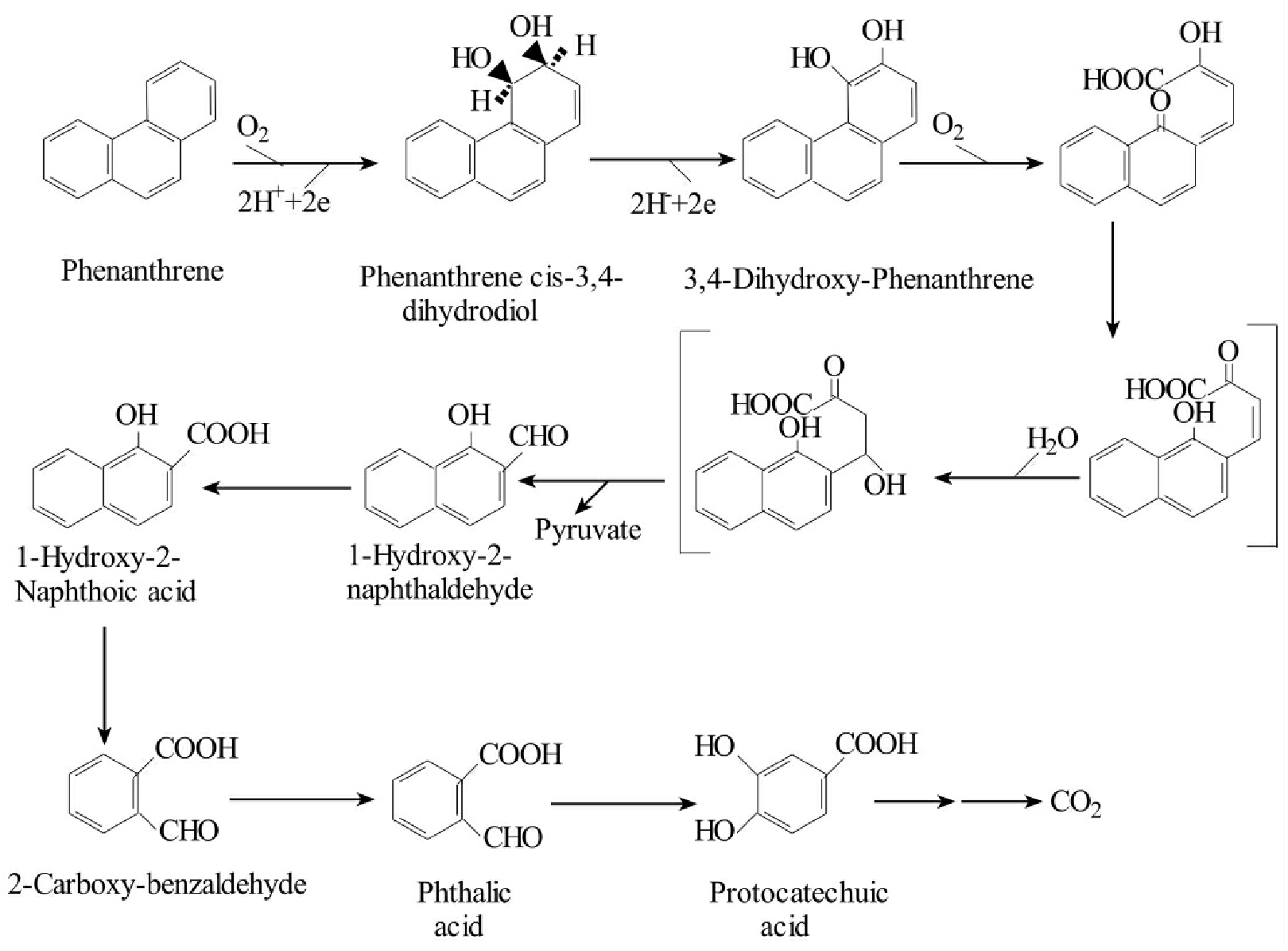
图8-27 假单胞菌代谢菲途径

图8-28 分枝杆菌菌株PyR1代谢芘的途径
四环的芘(pyrene)可被分枝杆菌降解产生CO2,中间代谢产物包括芘顺式-4,5二氢二醇(pyrene cis-4,5-dihydrodiol),4-羟基周萘酮(4-hydroxyperinaphthenone),4-菲苯酸(4-phenanthroic acid)、苯二甲酸(phthalic acid)和肉桂酸(cinnamic acid)以及反式-二氢二醇(trans-dihydrodiol)。还有其他的途径已被提出,这些途径综合起来如图8-28所示。相应的是红球菌属细菌对芘的代谢有不同的模式,包括1,2-和4,5-二羟芘(1,2-和4,5-dihydroxy pyrene),cis-2-hydroxy-3-(perinaphthenone-9-y1)propenic acid和2-hydroxy-2-(phenanthren-5-one-4-eny1)acetic acid。
在NADH存在时,甲烷氧化细菌荚膜球菌属的甲烷加单氧酶系统能氧化苯成酚,把萘氧化成1-和2-萘酚(图8-29)。酚再进一步降解。
许多种真菌、少数细菌和某些蓝细菌能产生细胞色素P450单加氧酶。这些酶能转化PAHS成为arene oxides,然后其被epoxide hydrolase水化(hydrated)形成trans-dihydrodiols或非酶促作用重排形成酚。只具有这些途径的微生物能够消除这些化合物的毒性,但不能利用其作为碳源。
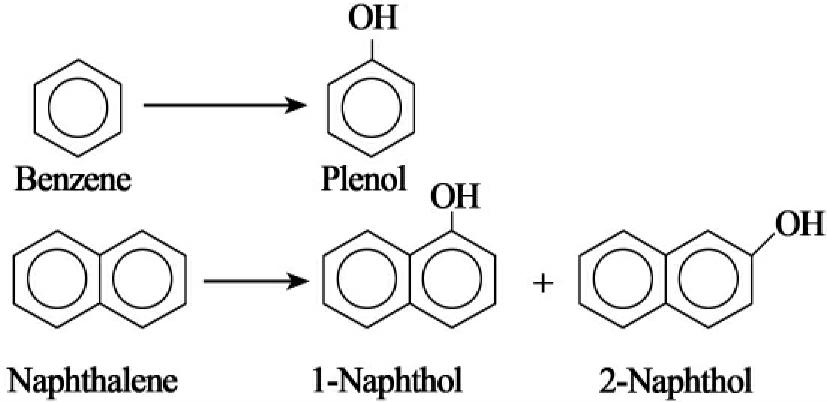
图8-29 甲烷氧化细菌氧化苯和萘途径
许多真菌能代谢二环萘成萘反式-1,2-二氢二醇(trans-1,2-dihydrodiol)、1-和2-naphthol、4-hydroxy-1-tetralone,以及glucuronide和sulfate鳌合物。包括蜡状芽孢杆菌、灰色链霉菌、分枝杆菌等某些革兰氏阳性细菌能代谢萘成1-naphthol。有的分枝杆菌菌株也能代谢萘产生trans-1,2-dihydrodiol。海洋蓝细菌某些颤藻也能代谢萘成1-naphthol。
菲是重要的三环芳烃化合物,能被众多真菌所代谢。雅致小克银汉霉(Cunninghamella elegans)代谢菲形成菲trans-1,2-,trans-3,4-和trans-9,10-dihydrodiols和一种糖苷复合物(glucoside conjugate)(图8-30)。其他真菌总状共头霉(Syncephalastrum racemosum)、白腐真菌原毛平革菌(Phanerochaete chrysosporium)、丝状细菌黄微绿链霉菌(Streptonyces flavovirens)、分枝杆菌的菌株以及海洋蓝细菌Agmenellumquadruplicatum也具有类似的代谢能力。
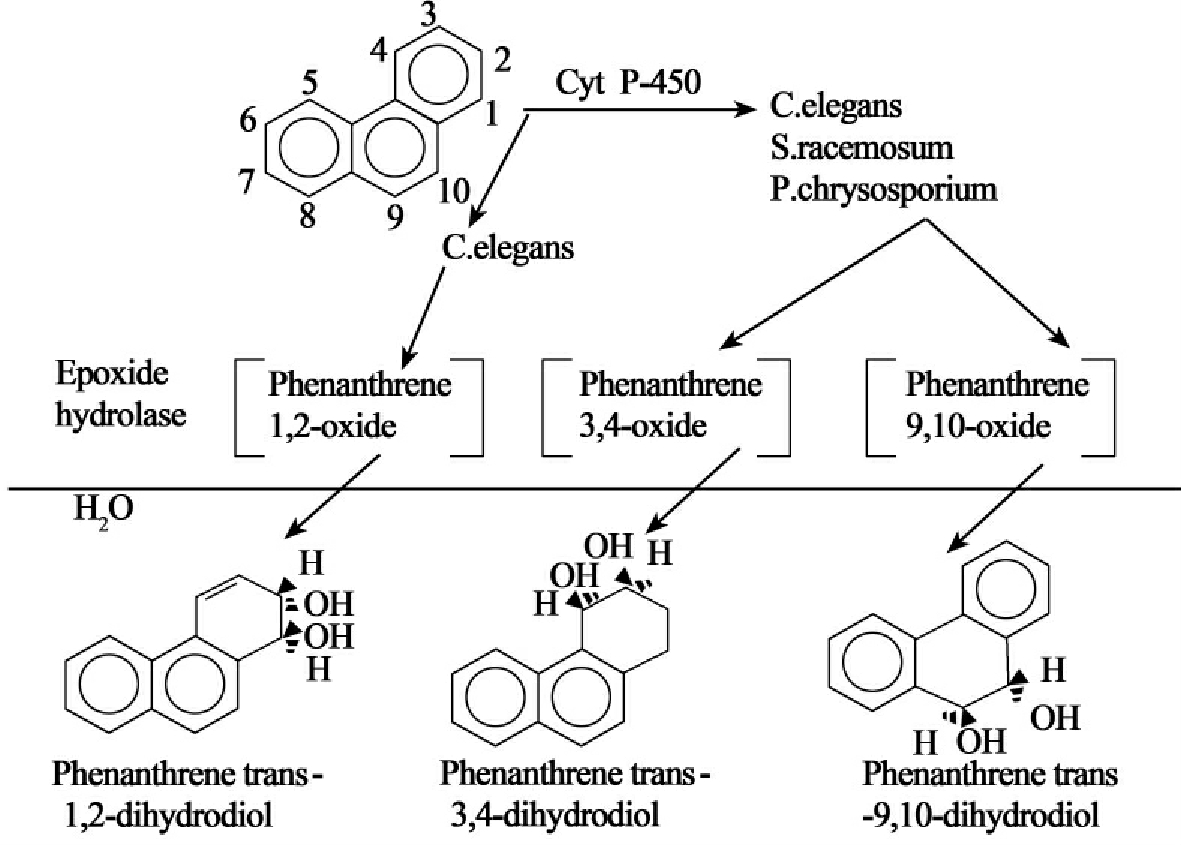
图8-30 不同真菌种对菲的代谢
(2)有取代基芳香烃化合物
有取代基芳香烃化合物特别是氯代芳烃化合物是一类非常重要,有广泛应用价值的化合物。它们被广泛用作溶剂、熏蒸剂(如二氯代苯)、木材防腐剂(如五氯苯酚)以及用作农药(如2,4-D、DDT、2,4-5-T等)。微生物降解这类氯代有机物的困难在于其碳-氯键非常有力,要使它断裂需要很大的能量投入。没有取代基的芳香烃化合物常见的中间产物是顺二羟基苯或儿茶酚。这需要紧邻的碳原子未被取代,而氯取代基能阻塞这些位置,增加生物降解的困难。甲基化的芳香烃的生物降解要么是攻击甲基,要么直接攻击苯环。烷基化的衍生物首先被攻击的是烷基的碳链,碳链经β-氧化后依碳原子数目的不同生成相应的苯甲酸或苯乙酸。然后是苯环的羟基化作用和开环。
苯酚和甲酚也是简单的带取代基的苯类衍生物,它们的降解途径如图8-31所示。
氯代芳香烃化合物是最常见的带取代基的芳烃化合物。二氯代苯和五氯苯酚是常见的氯代芳香烃化合物,它们的降解途径如图8-32所示。
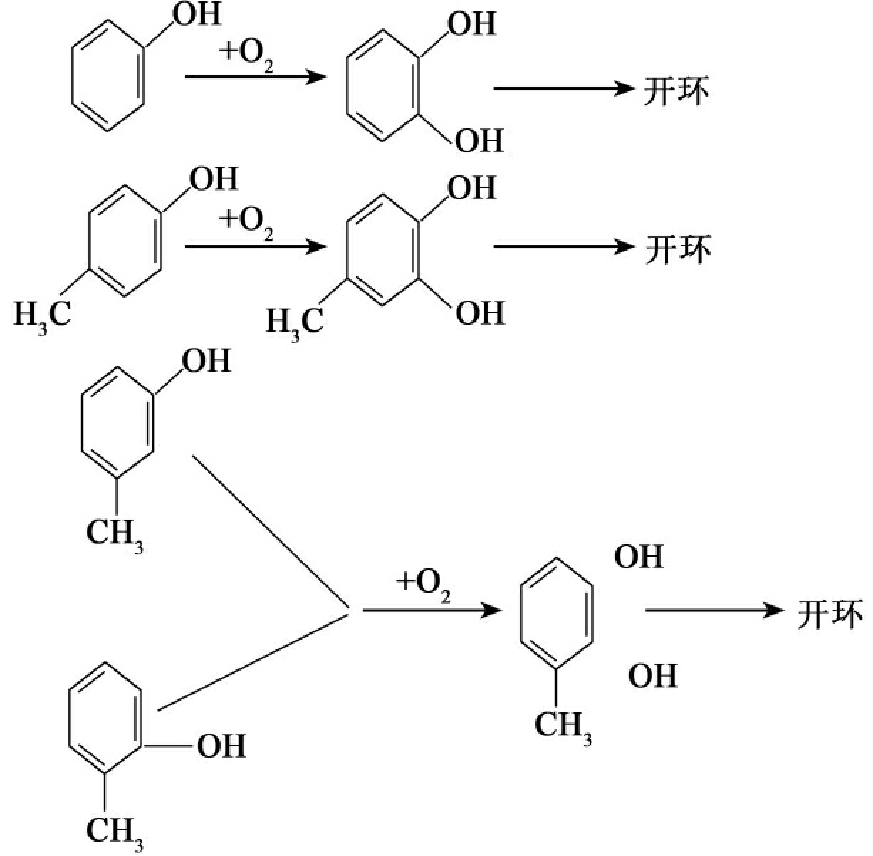
图8-31 苯酚和甲酚的好氧生物降解途径
2,4-D(2,4-二氯酚乙酸2,4-dichlorophenoxy acetic acid)是一种被广泛使用的农药。许多降解2,4-D的菌株已从全世界的各地分离出来,其中真养产碱菌(Alcaligenes autrophus)JMP134及其降解质粒PJP4被深入研究。其对2,4-D的降解途径在含酸有机氯化合物中具有代表性,特别是其对2,4-D的降解分别是由降解质粒和染色体联合编码的。最终降解过程导致琥珀酸的形成,氯被脱除。琥珀酸是一种生化中间代谢产物,进入到中央代谢途径,以产生CO2和H2或掺入到微生物生物量。能降解2,4-D的土壤细菌除产碱菌外,还包括节杆菌属、假单胞菌属、黄杆菌属、伯克霍尔德氏菌属(Burkholderia)、红育菌属(Rhadoferax)、不动杆菌属、棒杆菌属(Corynebacterium)、红假单胞菌属和鞘氨醇单胞菌属(Sphingomonas)。
2,4,5-T是和2,4-D结构类似的氯代化合物,其生物降解的速率大大慢于2,4-D,其降解途径也类似于2,4-D,其主要的差异在于多一次脱氯过程(图8-33)。
大量研究表明许多氯代芳烃化合物在厌氧下更易于生物降解,特别是还原脱氯是许多氯代化合物在厌氧条件首先发生的降解过程。五氯酚在厌氧条件的降解过程如图8-34所示。
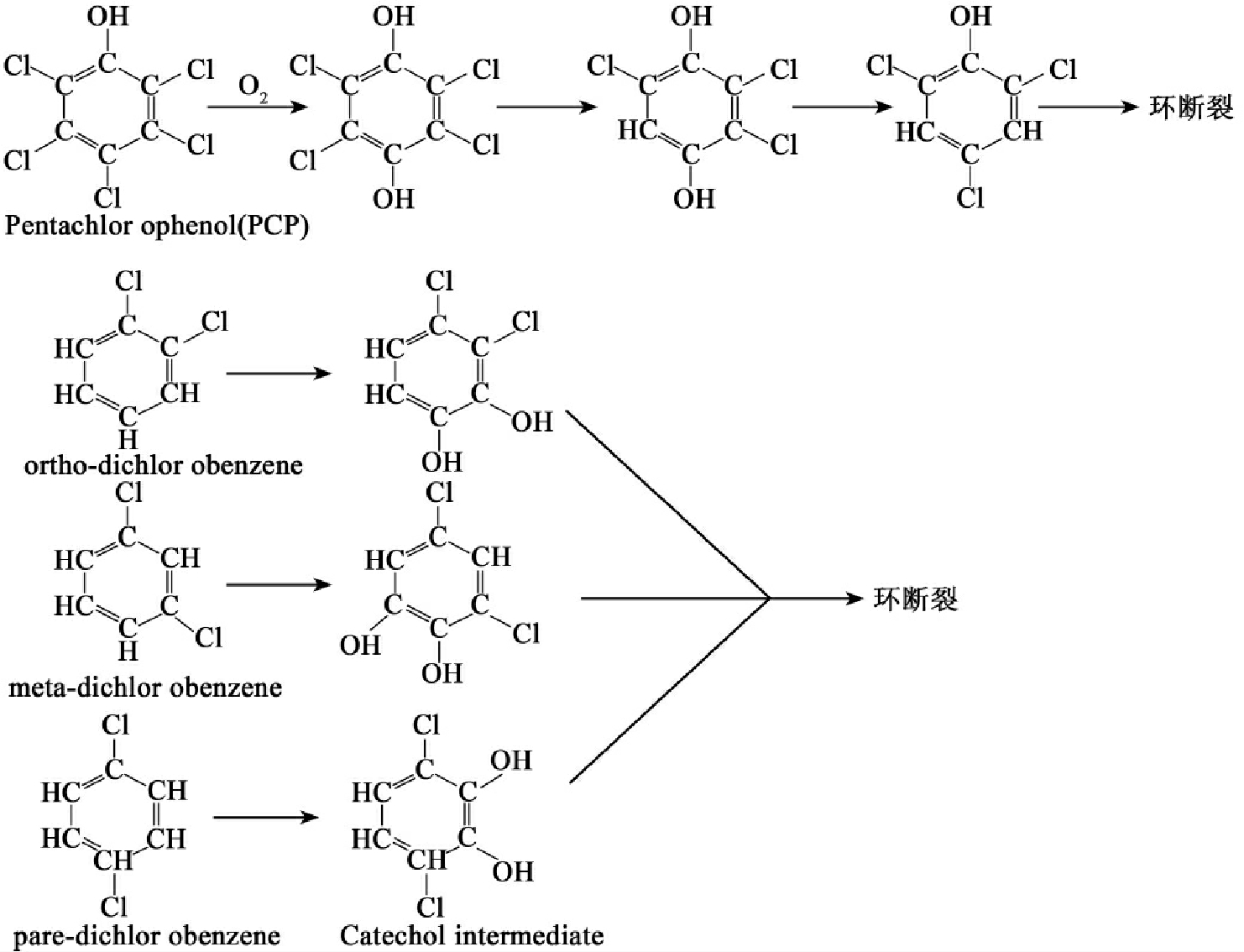
图8-32 五氯苯酚(PCP)和三种二氯苯最开始的好氧降解
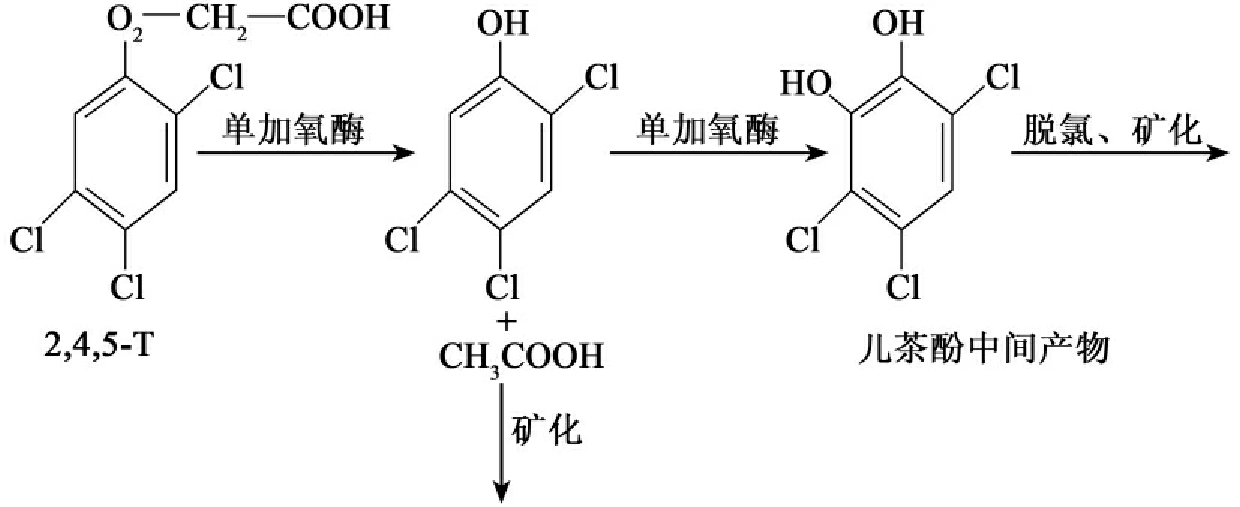
图8-33 2,4,5-T的生物降解
二、其他化合物的生物降解
其他化合物主要指有机氮、有机磷化合物。有机氮化合物可以作为微生物的唯一氮源。而有机磷化合物被降解后可以释放出大量的磷到环境中。
阿特拉津(atrazine)是一种在过去30年中一直被广泛使用的除草剂。已分离出多种能以这种除草剂为唯一氮源的微生物,其中假单胞菌ADP菌株的降解能力被广泛研究,其所带质粒pADP-1编码的酶可把阿特拉津降解成氰尿酸(cyanuric acid),而染色体上的基因编码的酶可以使降解产物进一步降解。阿特拉津的降解过程如图8-35所示。反应第一步是基因atsA编码组成型表达的阿特拉津氯水解酶催化完成的,反应中水分子的羟基置换三唪环上的氯,产物是羟化阿特拉津,并释放出盐酸。羟化阿特拉津的N-乙基侧链接着被atzB编码的氨水解酶水解而得到N-isopropylammelide。这种化合物再一次水解脱氨而得到氰尿酸,催化降解反应的是由atzC编码的水解酶。
尼龙(nylon)是纺织工业上应用广泛的多聚物,其基本单元是6-aminohexanoate。黄杆菌菌株K11225所带质粒pOAD2编码的三种酶能催化短的尼龙低聚物(2~20共价结合分子)生成6-aminohexanoate。这些酶都可以打断酰胺键,但攻击的是不同类型的多聚物。nylc的基因产物endo-型6-aminohexanoate低聚物水解酶能够线性化和解聚N-carbobenoxy-6-amin-ohexanoate三聚物。6-aminohexanoate环状二聚物的降解如图8-36所示。nylA编码6-aminohexanoate环状二聚物水解酶打开酰胺键中的一个,使二聚物线性化。nylB编码的另一种水解酶,能水解上述的降解产物存在的一个酰胺键,而后得到二个分子的6-aminohexanoate。
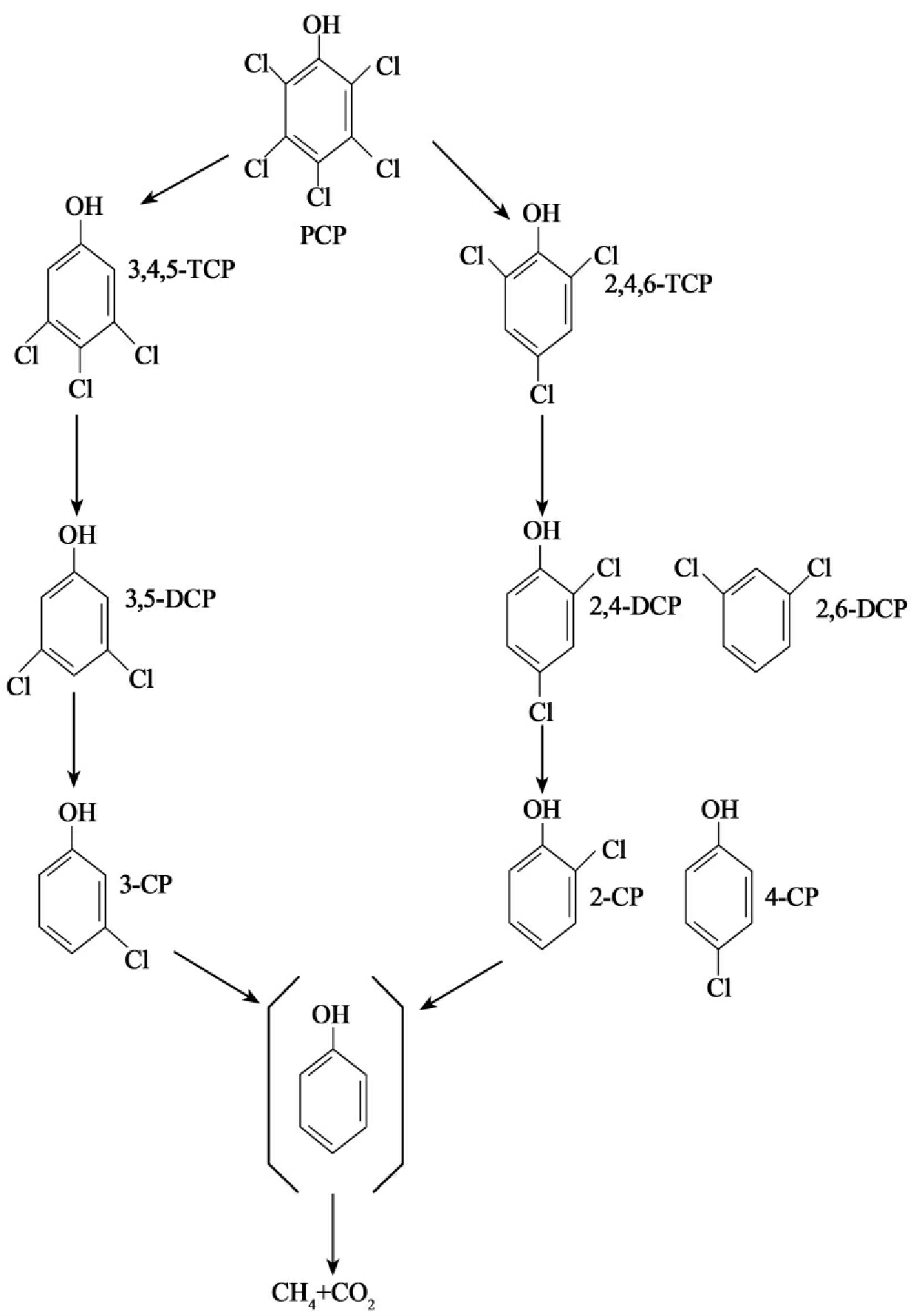
图8-34 五氯酚(PCP)厌氧条件下的生物降解
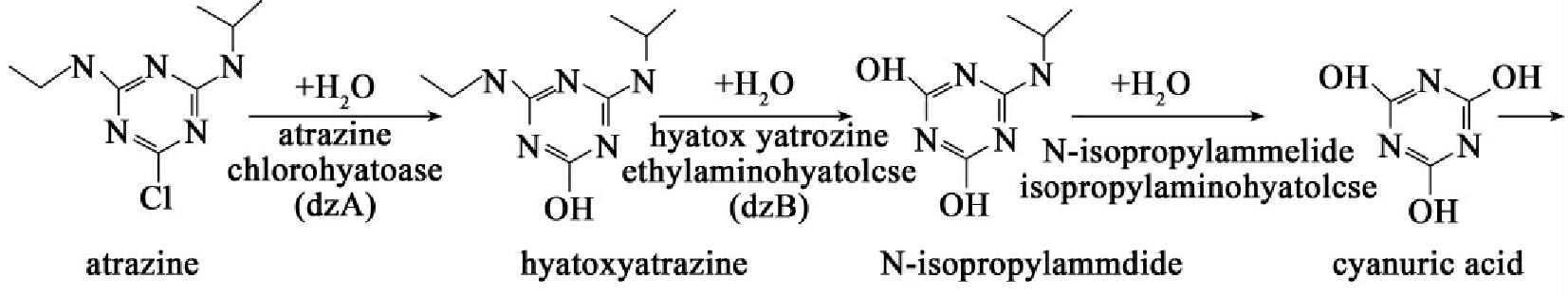
图8-35 PADP-1质粒编码酶降解阿特拉津途径
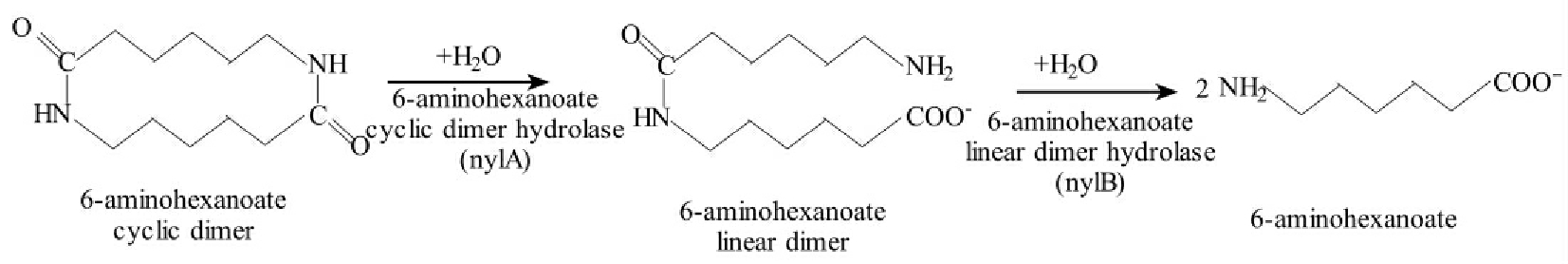
图8-36 pOAD2质粒上基因主导的尼龙低聚物的降解途径
对硫磷(parathion)是广泛使用的有机磷农药之一,常用于农业害虫的控制。此外许多有机磷类化合物还被作为化学武器。缺陷短波单胞菌(Brevundimonas diminuta)MG带有质粒pMCSI,质粒上的基因编码的广谱(broad-spectrum)有机磷水解酶打开对硫磷的磷酯键(phosphotriester bonds)。这种反应是水分子亲核加成到酐键(anhydride bond)的结果,得到二乙基(diethy1)thiophosphate和P-nitropheno1(图8-37)。opd编码的酶是组成型表达的,酶降解产物可被进一步降解。
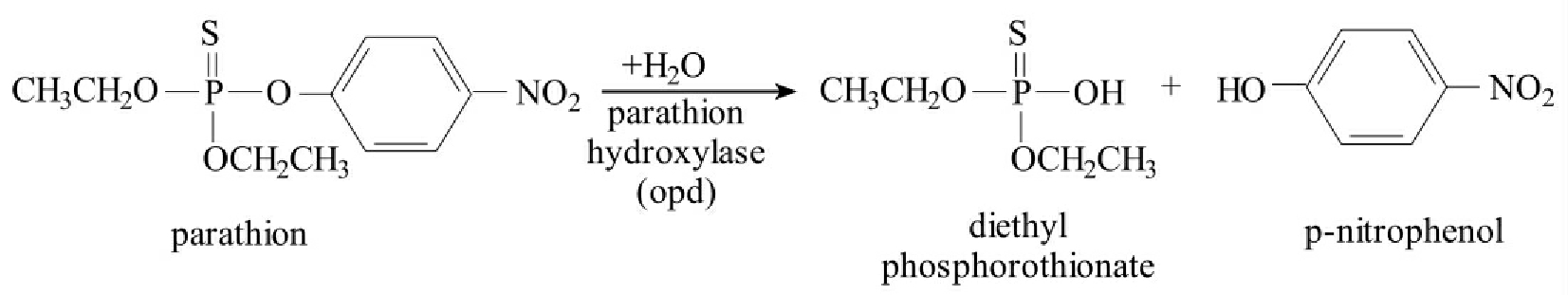
图8-37 质粒pMCS1质粒上基因主导的对硫磷的降解途径
第八节 有机污染物的降解性测定及归宿评价
种类繁多的各种化学物总会通过不同途径进入环境系统,大气、土壤、地下水和地表水等,它们的迁移能力和最后的归宿决定着它们将到达(分配)的位置,以及在这些位置中存在的浓度及时间。制约迁移归宿的主要因素是它们在环境中的行为,这些过程包括稀释、挥发、吸附、化学降解和生物降解,对化合物的迁移、归宿产生重要的影响。生物降解一般比化学降解更快、更完全,而且可以发生在非常广泛的环境条件下,生物降解使化合物的完整性和功能性丧失,并使化合物的组成成分回归到自然的无机物的循环中去。对一种化合物在使用前以及使用后进行生物降解和归宿作准确评价是环境保护中的重要工作。
一、生物降解性测定
1.测定方法
研究生物降解性的最根本的目的是依据生物降解的测定结果指示污染物的生物降解性,评估含污染物污水、废弃物生物处理的可行性和修复潜力以及它们在环境中的残留及生态风险。环境污染物的生物降解是一个十分复杂的过程,研究者可以按需要选择不同的终点,采用不同的方法来测定环境污染物的生物降解性。
(1)生物降解性测定的要素及表征
测定一种化合物的生物降解性,构建实际测定系统除了要有目标化合物外,必须充分考虑四个方面的要素:①降解微生物及其对污染物的可接受性,接种微生物可以有单种、菌群及混合菌群,微生物可以是专一性降解菌、适应驯化菌,以及污水处理厂污泥、污染环境微生物菌群、生态环境中微生物群聚生境(如底泥、土壤等)的微生物源。②降解系统的组成,从单一污染物加上其他营养物的培养液、污染物加环境样品模拟系统到微宇宙、中宇宙,甚至原位测定的野外现场调查。③检测终点,包括母体化合物的消失、矿化产物的产生(如CO2、NO3-、CH4等)、电子受体的消耗(如O2、NO3-等)、挥发性物质产生、能量产生(如ATP)、生物量增加、降解酶活性、生物毒性(母体化合物及中间代谢产物的毒性)等。④实际测定的环境条件,如pH值、温度、大气压、水活度、静态、动态等。
表征生物降解性的数据包括一级或二级速率常数、半氏常数、生物降解比速率(每克初始微生物,每小时对基质的去除数量)、达到标准化合物同样生物降解程度所需时间、BOD在理论需氧量中的比例、降解半衰期等。
(2)生物降解性测定的方法
任何生物降解系统都是目标化合物和上述四种要素的组合,一般来说都是一种模拟试验。因此实际测试中我们应根据研究目标设定不同研究方法。目前对测试方法的分类及命名都不甚统一,一般都是在满足其他条件下从某一角度出发设定具体的测定方法。
1)基于降解系统组成的测定方法。从降解微生物选择及降解环境系统来说有微生物方法和环境学方法。微生物学方法:其通常使用纯培养在最适条件下研究化合物的降解,然而其条件是自然环境所没有的,因此其结果不能直接预测它们在环境中的实际行为,降解性通常被高估,但对进行生物处理仍有重要参考价值。但使用的不同微生物接种物如在数量、驯化、纯培养、混合菌群、菌群等方面的差异可以得到不同的结果。环境学方法:其着眼于化合物在受污染水体和土壤中的降解性,通常使用取自污染区域或废水处理厂的混合微生物源或模拟自然条件下培养于实验室的混合微生物培养物来进行实验研究,对所得结果的评价更接近于野外的实际情况。模拟条件的不同情况使环境学方法实际上是极其多样的。
2)基于终点的测定方法。选择不同的测定终点也代表不同的方法,常用的有母体化合物的消失、O2消耗测定、CO2产生测定、活性污泥挥发性物质产生测定、酶活力测定、ATP量测定、总有机碳测定等。
①母体化合物的消失测定。母体化合物的消失是生物降解和生物转化的最具体最直接证据,对许多难降解环境污染物的生物降解性大多以母体化合物的消失作为表征,这是因为难降解污染物最初(或是第一步)的降解是极为关键的,开始的降解可以带动整个降解过程。这种方法也有其不足,母体化合物的消失并不等同于完全降解,还要考虑到中间代谢产物的抗降解、积累及毒性等问题。
②氧消耗测定。O2是好氧生物降解的电子受体,生物降解过程伴随着O2的消耗和CO2的产生,BOD测定就是建立在这种理论基础上。通过O2的消耗来判定生物降解性是一种十分科学的方法。O2消耗主要用华氏呼吸仪或电化学方法测定。通过测定氧的消耗测定基质的可生物氧化率、基质的生化呼吸曲线。
基质的可生物氧化率测定:基质的可生物氧化率测定以微生物作用下分解特定污染物的耗氧量为分子,完全彻底氧化所消耗的理论需氧量为分母,二者的比值即为基质的氧化率。
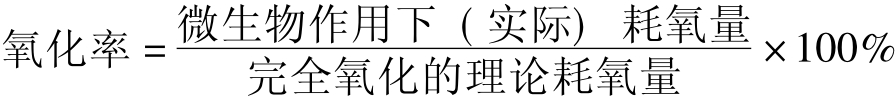
氧化率是可降解性的一种指标,氧化率越大,基质的可生物降解性越高。
这种方法在实际的应用中可以用COD或TOD替代完全氧化的需要量,以BOD5替代实际耗氧量,以BOD5/COD/或BOD5/TOD比值可评定其降解性。比值越大,说明该物质越易生物降解。据国内外有关研究报道,基质氧化率和生物降解性的关系如表8-11所示。
表8-11 氧化率与生物降解性

基质的生化呼吸曲线测定:测定基质降解过程中的耗氧量,以时间为横坐标,耗氧量为纵坐标,即可绘制出基质的生化呼吸曲线,亦称耗氧曲线。当微生物处于内源呼吸阶段(即利用自身细胞物质作为呼吸基质)时,其呼吸速度是恒定的。耗氧量与时间成直线关系,这条线被称为内源呼吸线。当供给微生物有机营养时,耗氧量随时间的变化是一条特征曲线,称为生化呼吸线。为评价基质的可生物降解性,常将基质的生化呼吸曲线与内源呼吸线进行比较。该比较可以出现如图8-38所示的三种情况。
③脱氢酶活性测定。降解酶在生物降解中起重要作用。脱氢过程在微生物氧化分解有机物中起重要作用。有机物的生物降解性能可以从脱氢酶的活性上明显反映出来。脱氢酶对毒物尤其敏感,它的活力下降往往与毒物的侵袭有关。因此,测定脱氢酶的活性也是进行有机物生物降解研究的常用方法之一。
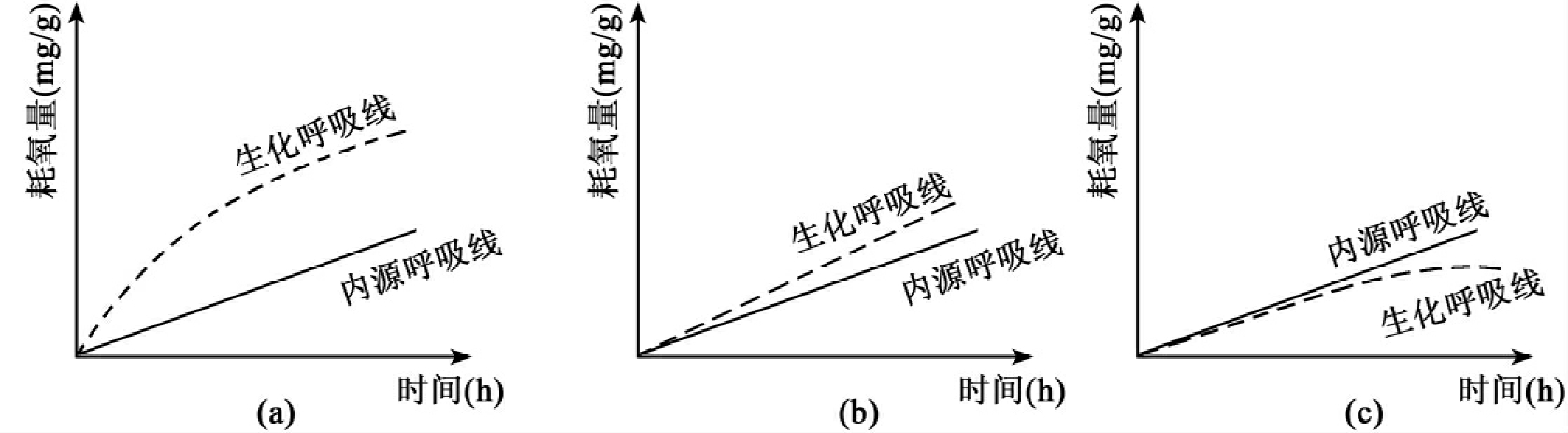
图8-38 微生物呼吸线与内源呼吸线的比较
a.生化呼吸曲线位于内源呼吸线之上,说明该有机物可被微生物氧化降解,两条呼吸线之间距离越大,说明该化合物降解性越好。b.两条线基本重合:说明该有机物不能被微生物氧化分解,因为虽然投加了基质,微生物所进行的仍只是内源呼吸,但它对微生物的生命活动亦无抑制作用。c.生化呼吸曲线位于内源呼吸线之下:说明该有机物不仅难以生物降解,而且对微生物产生明显的毒害(或抑制)作用。生化呼吸曲线越接近横坐标,表明毒害越大。
④ATP量测定。三磷酸腺苷(ATP)是生物体内一种高能的磷酸化合物,它在能量的贮存和转换中起着重要的作用。测定ATP的含量,不仅可以反映生物的活性,而且还能反映活性生物量的多少。目前已有快速测定ATP量的精密仪器,故这一方法已越来越引起人们的注意。
⑤总有机碳(TOC)测定。取一定量样品经微生物作用,然后测定作用前后试样中总有机碳的变化,以此来评定被测有机物样品的生物可降解性。有机物所含有的总碳量叫做TOC(total organic carbon)。水中的碳除了包含于有机物中外,还存在于碳酸、重碳酸中。因此,由总碳量TC(total carbon)减去无机碳TIC(total inorganic carbon)便可求出TOC。测定时把微量水样通过温度达到950℃的燃烧炉,使水样中的有机碳和无机碳全部氧化,生成CO2后用紫外线气体分析仪测出;与此同时,将另一份同样的水样在150℃左右的温度下,使无机碳化物氧化,生成CO2并测定之,二者之差即为TOC。
因为有机物的生物降解也可看做是含碳有机物物质的无机化和转变为细胞物质的过程。所以只要掌握好一定的活性污泥和时间等条件,所得到的检测试样中总有机碳的变化值,便可用来表示相应有机物的可生物降解的程度。
⑥CO2产生量测定。有机碳化合物完全矿化降解的最终产物是CO2,测定降解过程中CO2的产生量是对生物降解中有机物转化成CO2程度的最好指标,尤其适于监测混合有机物的降解过程。同时CO2收集和测定简便精确,因此测定CO2来监测生物降解过程特别重要,例如监测堆肥过程中CO2的产生量可以有效指示处理过程及效率。
⑦活性污泥中挥发性物质测定。活性污泥中的挥发性物质,是活性污泥中具有生物活性的部分。因此,活性污泥中挥发性物质含量的多少,可反映一定条件下生物活性的强弱。生物活性可间接表示基质的生物降解性能,因而假如其他条件保持恒定,仅仅改变待测物质的种类或浓度,那么经过一定时间后测定活性污泥中挥发性物质的含量,所得数据应与有机物的生物可降解性有关。
⑧专一性14CO2测定。收集测定14C标记有机污染物生物降解放出的14CO2,并以此评价这些有机污染物的生物降解性具有科学、特异和精确的特点,是生物降解测定中首推的方法。这种试验分批进行,放射性标记试验化合物和实际的环境样品一起培养,监测放射性产生能力。这种专一性使我们能测出一种化合物的真实浓度,证明生物降解的完全性,以及实际的动力学过程。这种方法已被用于活性污泥、河流河口、海洋水体、淡水和海洋沉积物、地下水和表层及表层下土壤降解。这种试验方法有许多优点,首先其易于测定,CO2易于吸收在碱性溶液中,也易于通过液体闪烁计(LSC liquid scintillation counting)定量测定。这种方法也有某些限制,用CO2作为唯一的终点,只能确定化合物经历矿化以后的归宿,不是矿化而是转化的化合物有较少的应用,而且有时也不与母体化合物的消失相适应,此外化合物及其代谢产物被综合到生物量或自然腐殖质材料中而造成在某些系统中的低回收使得某些物质的归宿带有不确定性。这种低回收的情况难以测定单个化合物在复合物中的生物降解程度及存在的代谢产物,而且这种唯一一种终点分析使我们对降解原理的理解不足。
3)基于有机物降解难易的测定
①易于生物降解化合物的降解试验。易生物降解物质应满足下列条件:
a.它们能作为微生物的唯一碳源与能源。
b.它们能完全矿化成CO2和水。
c.它们不需要任何复杂的适应和选择的微生物降解菌。
d.它们的降解速率和存在于污水中的有机物相应。
e.降解快速进行,且不出现不希望的环境效应。
模拟自然条件用相应浓度的基质和仅部分适应的微生物,当试验已经证明易于生物降解时,则能确实地假设这种化合物在自然条件下也易于生物降解,不需要进一步试验。试验通常包括BOD分析和CO2产生来决定TOC的失去。但阴性结果不一定说明这种化合物在各种环境条件下是抗降解的。
②潜在生物降解性化合物潜在降解性测定。其目的是决定是否降解是基本可能的。潜在降解性试验一定程度上模拟生物污水处理厂和自然条件,维持一定的条件,为微生物降解提供基本条件。在这种条件下不能充分达到希望降解程度的化合物被定为难降解的(poorly biodegradable)。这些条件的限定是非常重要的,分类为难降解的基质的降解在环境中已被证明很大程度上依赖于具有长世代时间的微生物。用生物柱和生物膜反应器进行实验倾向于能富集更长世代的微生物。这就是说降解基质时要用选择的微生物,使用常用的污水处理过程的降解试验不能说明易于生物降解。
③厌氧条件下的生物降解试验。大部分有机物在好氧条件下更好转化,但有些化合物在厌氧条件下能更快降解。例如许多卤代化合物(特别是多氯联苯等氯代化合物)在厌氧条件下能更好脱氯。厌氧生物降解性评价有特别重要性,因为污水处理中污泥的修复在厌氧条件下进行,某些工业污水在厌氧过程中成功处理,在自然条件下,厌氧降解可以发生在河流、湖泊沉积物下层及地下水的深水层。厌氧过程在试验类型中应有其重要地位,特别是那些在好氧条件下具有抗性的化合物。
④降解试验层次性方法。首先进行易生物降解试验,但一个单独的试验的正和负的结果对基质的降解性评价是不充分的,要多种试验并行,通常包括结合BOD分析和CO2产生来决定DOC失去。如果两种试验证明易于生物降解,这种试验即告完成。如果试验结果是阴性的,接下来进行潜在生物降解性试验。如试验仍达不到希望的降解性程度,基质被定为难降解的(poorly biodegradable、resistant biodegradable)。一般来说使用特殊的试验方法,即要选择微生物和较不普遍的污水处理过程,则化合物不能认定为易于生物降解。如果需要,研究工作的下一步是对降解微生物进一步研究。
现存的许多生物降解试验方法主要存在的问题是不模拟野外的实际情况,局限于选择的终点,所确定的化合物浓度多从实验出发有利于实验而不是考虑化合物正常进入或存在于环境基质中的量。此外动力学分析常不进行,而且数据先被套入一种预先设定的模型(如一级反应)。
用不同的方法测定的化合物的降解性其总的趋势是一致的,但具体的数值因不同条件的差异有较大的差异也是正常的。
2.实验设计
(1)试验化合物的剂量和加入
实验设计对生物降解(包括实际的14C试验系统)有重要的影响。设计中的一个重要方面是化合物的剂量及如何被加到系统中。合适的剂量应是环境中的实际浓度,但试验研究中的剂量往往高于环境浓度,因此提高测试灵敏度是一个值得关注的问题。有研究者对一种与日常使用阳离子表面活性剂相对应的长链胺化合物在活性污泥中的矿化作了研究,化合物采用两种不同的处理方式(分别溶于异丙醇和20%异丙醇-水两种系统中)被注射到试验系统,结果发现矿化的差异非常明显,前者的矿化快而彻底,后者却慢而不完全,如图8-39所示。
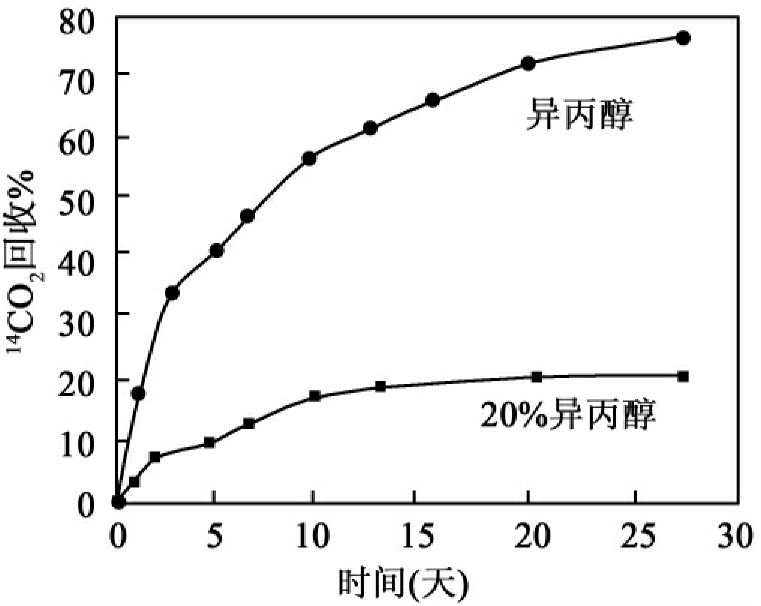
图8-39 长链胺化合物在活性污泥中的降解时剂量的效应
一种硬脂酸被以水溶液的形式注射到土壤,其在土壤的不同组分中的矿化也充分说明矿化速率的剂量效应(图8-40)。
从图中可见其在沙中矿化最快,而在棕黄酸中矿化最慢。这些例子说明矿化速率和矿化程度高度受到化合物的剂量及被接受试验系统的影响。产生不同结果所提出的问题是什么代表真实,什么是正确的。在第一个例子中两种注入方式都不能准确代表这种化合物进入废水的方式。但这种来源于注入方法的生物降解试验敏感性说明一种化合物在环境中的物理/化学形式的关键重要性,而且需要在实验中再现这种形式。
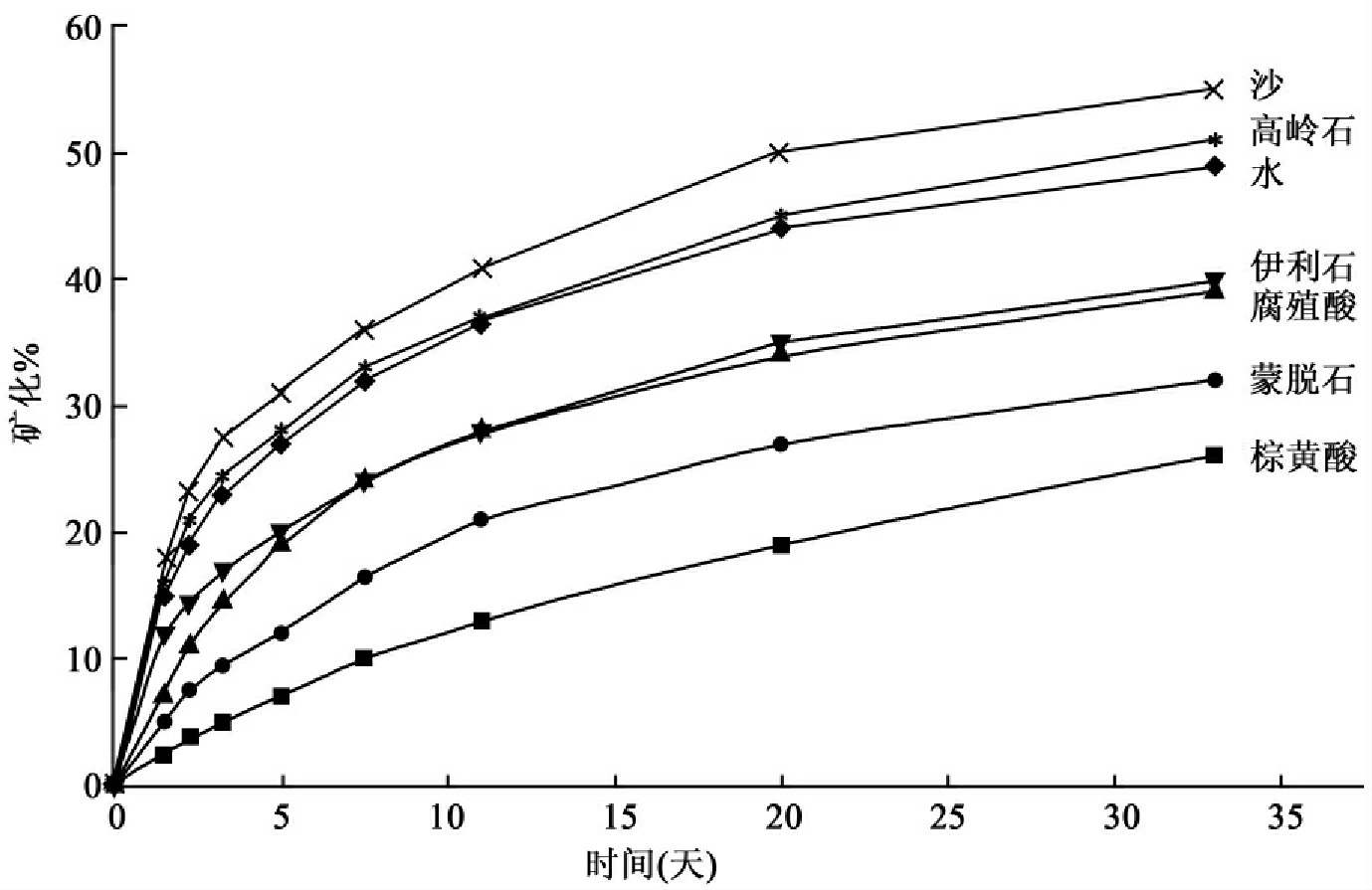
图8-40 硬脂酸钠在土壤中矿化时剂量的效应
(2)数据处理
数据处理是实验设计中一个重要方面。现在的问题是常常没有进行动力学分析或数据被局限于事先认定的模式中,而一般都用一级反应的模式来描述生物降解和化合物的消失。有人研究过一种阳离子表面活性剂(AES,alkl ethoxylate sulfate)的生物降解,经拟合AES的初级降解可用一级衰减模型描述,AES的半衰期少于5分钟,但实际检测数据说明当模型指示没有AES的30分钟后仍存在,实际的低水平(<1%)仍然检测到(即使在3小时后) AES。利用一种简单的一级消失模式和几分钟的半衰期会明显高估污水处理的去除。最适合的方法应是按准确测定的数据拟合出降解动力学数学模型。
3.生物降解试验方法改进
理想的生物降解试验应包括母体化合物消失的精确分析,代谢物的形成、消失及矿化。同时找到一种最合适的描述公式。改进生物降解测定方法主要包括三个方面:①增加检测的终点,包括母体化合物、代谢物和放出的CO2。②以一种更符合实际的方式把化合物加到试验系统。③对数据进行更强有力的动力学分析以准确测定生物降解速率和找到最合适的描述生物降解过程的模式。
(1)多终点检测
增加检测的终点包括母体和代谢物的量,特别要测定综合到生物量成分(即蛋白质、脂质、核酸和细胞壁)中的量。为此发展了一系列的分析技术,包括Rad-HPLC、Rad-TLC和Rad-GC/MS以及系列抽提方式回收母体、代谢物和生物量成分。
图8-41是这种多终点的试验流程,称为die-away试验。试验的化合物培养于新近从环境中得到的样品(如活性污泥、生污水、河水、河水稀释的排放水、厌氧消化池污泥等)。氯化汞灭菌的样品作为非生物对照,这种处理作为分析母体化合物回收、各种生物量组分放射性的非专一性回收及水解、吸附在试验容器或挥发的非生物失去的对照。定期从两种处理取子样品和冻干处理,冻干固体用合适的溶剂抽提以回收类似代谢物,抽提物用LSC分析测定总的放射性,用Rad-TLC测定相关的母体和各种代谢物的丰富度。提抽后的固体被直接分析或定量转移到微离心管作生物量的生物化学分馏。分别用冷三氯乙酸抽提以回收低分子胞质成分,用醇/醚回收脂肪,热的TCA回收核酸,用10mol/L NaOH回收蛋白质。此后再以溶剂提抽并离心,并用LSC计数回收悬浮物的放射性,最后是全部抽提后固体焚烧测综合到细胞壁的量,每次分馏的数据与非生物对照比较。另外14CO2通过酸化子样品测定,并比较两种处理的不同。放出的14CO2用碱液回收并用LSC定量测定,溶解的14CO2通过酸化子样品测定。吸收酸化后放出的CO2并在碱液中测定。通过比较两种试验的差异我们可以理解化合物在环境样品中的生物降解情况。
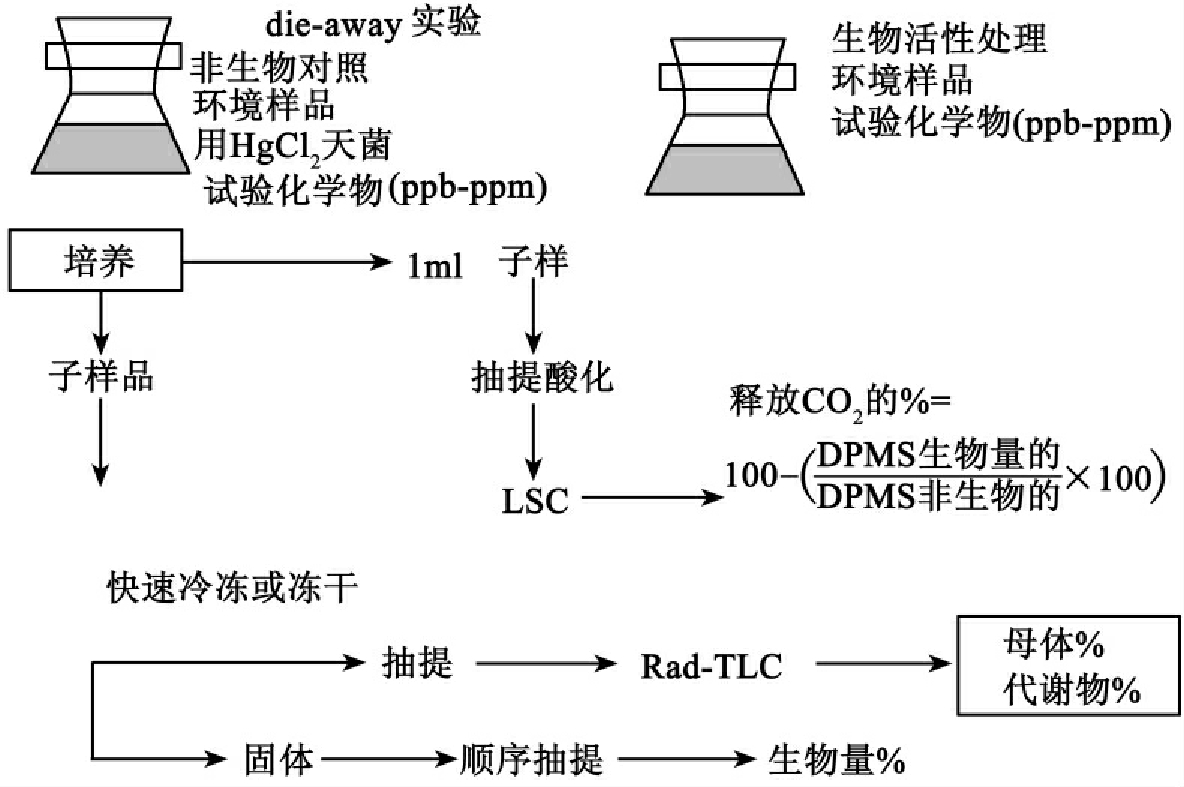
图8-41 普通die-away试验的示意图,用于评价14C试验化合物在环境样品中的初步和最终生物降解
图8-42是这种试验设计所得数据的图形。试验说明了AES(alkyl ethoxylate sulfate)在活性污泥中的降解,试验化合物是一种14CE3S同系物,均在1和3羟乙基上标记,注入剂量最后浓度为1mg/L。从图中可见化合物加入后化合物母体的消失立即开始并快速进行。和母体消失同时发生的是放射性被吸收到生物量中,14CO2的放出和极性代谢物的短时间(瞬时)出现,这种代谢物被鉴定为PEG磺酸盐(polyethylene glycol sulfate)。这一结果说明AES降解是在醚键处断开导致PEG磺酸盐和脂肪醇的释放,脂肪醇其后又被矿化和综合到生物量中。经过3小时的降解,大约95%放射性从母体中放出,这等同于在生物量和14CO2中的分配。这种分配和大约0.5的生长收获是相一致。连续试验更长的时间仅会得到生物量中的碳转化成14CO2的数据。从这组试验数据可以看到使用矿化作为唯一的终点将会总体上高估降解母体化合物及它们初级代谢产物所需要的时间(即把短的降解时间估计长了)。
(2)受试化合物的模拟注入
试验设计中分析以外的一个重要问题是把受试化合物注入到试验系统中。对中高溶解度化合物,将其水溶液注入即可,但对大多数低水溶性和吸附性化合物则要特别注意注入方式,实际测试中一般应模拟其进入系统的方式。例如某一种洗涤剂成分进入污水的过程是其和各种其他清洁剂,特别是与表面活性剂一块被排放到下水道,然后进入污水系统。由此接种这种化合物到试验系统的最好方式是把其溶入污水中。这种方法不仅模拟化合物如何正常进入污水,也使化合物在试验系统中的均质分布,还改进了取子样品时的可替换性。
又如研究洗涤剂在土壤中降解时,先要考虑其是如何到达土壤的,大部分的洗涤剂成分是作为污泥的一部分到达土壤的,而污泥是作为土壤调理剂(amendment of soils)使用的。试验化合物接入土壤的理想方法该是把少量的污泥样品置于试验容器(test vessel)中,以溶质或水的形式直接把试验化合物加入污泥中,把土壤加到容器,并混合污泥和试验化合物到土壤,因此这种方法可更加准确模拟洗涤剂成分如何正常进入土壤,而不是注入到土壤。这是一处模拟自然过程的方法。

图8-42 AES在活性污泥中开始降解的情况
(3)数据动力学分析
对数据的动力学分析也是试验中的重要问题,已有商业出售的回归软件(Jandel Table Cwrve2D)把数据输到多重等式(multiple equations),以进行非线性回归。实际上,每一生物降解数据系列能符合于各种衰减和利用等式。这些等式包括:零级(zero-order)、一级(first order)、统计学一级(logistic first-order),以及firse-order with log or three-helf order with growth or three half-order without growth。最合适模式可用统计学方法作检验。图8-43说明选用最适模型的必要性。从图中可见一级(first order)和three-half order模型符合于一种AES同系物在活性污泥中初步生物降解数据。r2检验说明一级(first order)衰减模型准确描述AES的消失,实际符合(fit)的closer的检测和残留物说明3/2-order模型更加合适和准确。
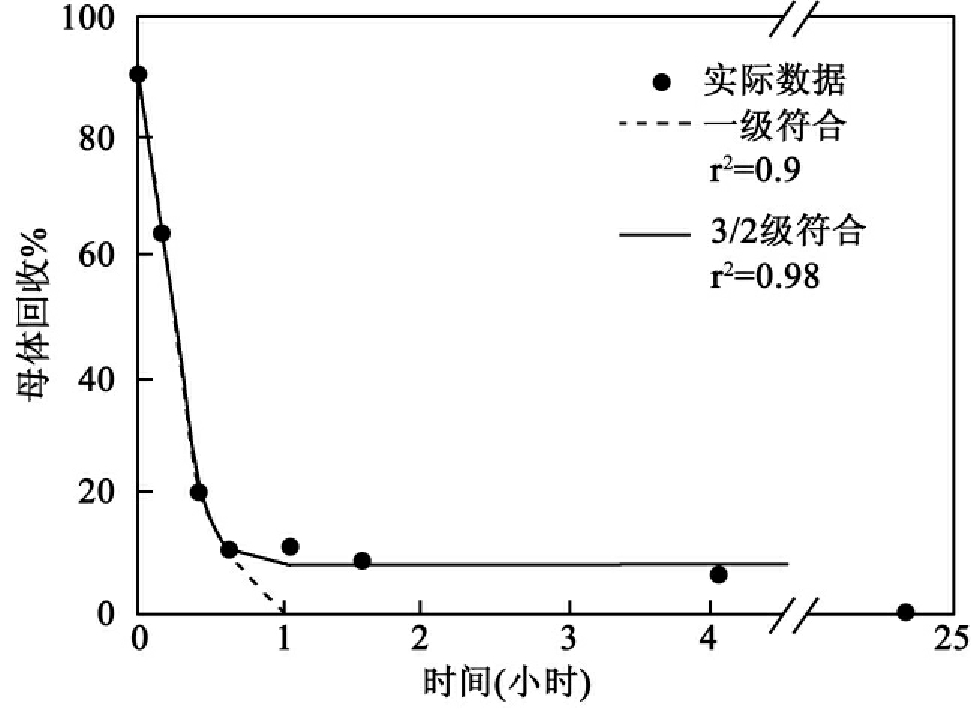
图8-43 一种AES同系物在活性污泥中的初步降解:一级与3/2级模型与实际数据符合比较
二、生物降解性评价
生物降解是环境中许多合成有机化合物消失的主要机理,生物降解速率是任何暴露模型的最重要参数。生物降解性评价一般要回答以下的问题:①化合物是完全降解还是部分降解?是易于生物降解,还是难以生物降解?难以生物降解化合物的降解速率。②化合物在环境中的半衰期。③有毒化合物毒性的消失状况。
生物降解评价中如果一种化合物能完全降解,则安全评价的焦点是母体化合物而不是任何化合物代谢产物的影响。降解在化合物存在的环境中能否实现,其生物降解过程的动力学是一个重要问题。对部分降解化合物关心的问题不仅是在生境中降解,而且还包括以一定速度降解是否足以降低暴露浓度,防止在整个时间内积累。部分生物降解也有真实的环境好处,这包括代谢产物在整个环境的较少分散和较低的环境的浓度,较低的浓度能转变成较高的安全系数。
为了进行科学的降解评价,需要得到可靠、精确的生物降解数据,这些数据最好在模拟原位的条件下取得。在测定生物降解动力学数据中,最重要的问题是测试化合物的浓度的确定,最科学的浓度应是化合物存在浓度。另外,化合物的环境组成应模拟原位的环境。在某些情况下,一种化合物的环境组成对降解的速度和程度有明显的影响。另一方面微生物群落的水平、组成和生理状态对原位发生的降解有重要影响。微生物群落的适应决定着一种可降解化合物降解的速率和程度,是否使用预先暴露或驯化群落取决于化合物进入环境的输入模式。如果一种化合物是偶然或不连续的释放,这时使用一种预先驯化的群落来测试可能产生错误的结果。如果一种化合物是连续排出,群落将得适应和继续适应,因为污染物对环境的污染代表一种新的生态位。在这种情况下,相应的生物降解速率适应新群落的速率。接种微生物生物量的高低也对降解产生重要影响,有人研究低微生物群体作为微生物源降解易降解的线性烷基苯磺酸钠盐时,发现其降解率经历30天后仍比用活性污泥作为微生物源低得多。为了避免使用过低的降解微生物生物量,有人建议接种物的内源呼吸在28天试验中不应低于0.5mg/L。最后试验基质将是相应的,如果要测定的是某一种化合物在活性污泥或土壤中的降解速率,试验应在活性污泥或土壤中进行。这种污泥和土壤具有所有与它们相关的贡献,包括固体、水量、pH值和氧浓度。
测定和筛选出的生物降解性数据必须小心评价。降解速率只有在所有的实验条件(特别是接种物)一致才能确实地进行比较。测定有机基质生物降解性的OECD方法允许使用广泛的微生物接种物(从来源和数量两个方面)。表水、处理厂排出物和活性污泥都可以作为接种物。接种物的量从每升基质数毫升到数十毫升,这样接种物的微生物数量相差上十倍。这样使用不合适的接种物来源和数量会造成假的负结果,因为它不仅能影响生物降解的总体过程,也影响降解程度。在这些情况下,实验室间实验结果比较是困难的。一个实验室把一种基质划到易生物降解,但另一个实验室可以把同样的化合物划到难生物降解。使用少量接种物(低量微生物)往往造成这样的问题。这种方式取得的数据和实际的降解性不一致,实际有机基质的降解性被低估。
三、建立在生物降解测试基础上的归宿评价
归宿是一种化合物进入环境经历一段时间以后的存留状态。归宿评价可以预测特定化合物对环境的影响。定量的归宿评价依据的是化合物的内在的物理化学特性,这包括挥发压(vapor pressure)、Henvy's定律常数、水溶性和Kow(辛醇水分配常数)以及QSBR(quantitative structure biodegradation relationship)预测或筛选水平生物降解试验(screening level biodegradation test)以及实际的生物降解测试。
生物降解测试给我们真实可靠的化合物的降解情况。建立在生物降解测试基础上的归宿评价比仅依赖物理化学特性以及QSBR要科学可靠得多。试验结果可以得到对一种化合物环境归宿的一种定量指示,说明其是否将分配到大气、土壤、沉积物或水柱,说明化合物的真实存在。当然在实际的归宿评价中只有生物降解的数据仍然是不够的,因为实际的测定并不能覆盖所有的实际情况,仍需要把化合物的各种参数、负荷、迁移等综合到评价的数学模型才能对环境污染物在环境中的归宿作出评价。需要指出的是许多归宿模型是不准确的,因为:①许多模型不能准确指示各种归宿过程和它们的相互作用。②数学模式对归宿过程的不适当表达。③模型的不适当参数化,如参数是在化合物不适当浓度、不适当基质的生物测试中得到的。
四、生物降解性测定的完整过程
生物降解的过程总体上和传统上的微生物纯培养是一致的,都是利用基质取得能量和前体物质合成新的细胞生物量,但又有明显的差异,这些差异表现在:①测定常在野外现场或用取自野外地点的样品(水、土、沉积物、工业排放水等)进行。②相关的基质是环境污染物,其和微生物生理学家传统的研究基质结构类似性较小。③被研究的污染物在环境中的浓度常常很低。④自然的微生物群落作为降解反应生物群,人们对它们是不了解的。⑤当用纯培养研究污染物代谢时,生物一般已从野外生境分离,而选择的依据是培养物的生物降解能力。
生物降解性测定从方法学上说应是设计出一系列实验来证实、测定和揭示污染物的纯化学改变以及对有机污染物归宿产生影响的相关的微生物生物学和遗传学特征。这个过程类似于证明传染病病原体的过程。完整的生物降解研究包括二个阶段(图8-44)。第一阶段处理土壤、沉积物、水和工业排出水样品,并构成一个黑箱(black boxes),再以分析化学方法测定污染化合物消失。第二阶段开始污染物降解微生物的分离纯化,然后再进行精确的生理和酶学评价,并对编码降解基因的DNA序列、表达及调控作分子水平的研究。但一般的生物降解试验仅进行第一阶段。
第一阶段:实验富集和净代谢活性的测定

第二阶段:纯培养的分离和污染物代谢的生理学、生物化学和分子基础的检测

图8-44 理解生物降解过程的二个阶段方法,第一阶段从环境样品开始,第二阶段延伸到单种微生物代谢污染物的生物化学和分子方面
第九节 有机物的结构与生物降解性及归宿预测评价
面对“寂静的春天”,化学家、环境学家、政府官员深刻反思,我们能否合成不使环境污染、不损害人体健康的化合物呢,我们能否在人工合成化合物进入环境前就了解它们在环境中的行为呢?科学家对此作出肯定的回答,并由此提出了绿色化学的观念,我们能够预知一种有机化合物的生物降解性,能够合成对环境友好,不损害人体健康的化合物。这样有机物化学结构与生物降解性的相互关系就成为生物降解中的一个最重要的问题。
一、有机物的结构与生物降解性
生物降解性(biodegradability)是说明基质被微生物过程改变接受性特性的概念。有机化合物的生物降解是它们分子结构的部分简单化或完全瓦解。大量的实验研究结果和实际的监测数据都雄辩地说明有机物的化学结构与生物降解性之间存在着密切的相互联系。
在长期的研究中我们积累了大量的有机化合物生物降解性数据和资料,许多人对其中的规律作了归纳总结。一般认为下面的分子特征是抗生物降解的。①卤素,特别是氯和氟的取代;②链分支,特别是季碳和叔碳;③硝基(nitro)、亚硝基(mitroso)、偶氮基(azo)、芳香胺基和arylamino基;④多环残基(如PAHS),特别是有3个以上的叠加环;⑤杂环残基,如嘧啶环;⑥脂肪烃醚键(aliphatic ether bonds)。
在大多数情况下这些特征影响一种化合物作为诱导剂基质的能力。例如把一个Cl原子加到苯环上就使环较不适于加氧酶的攻击。上述列出的肯定不是增强抗性的全部基团,而且一种化合物的抗性的产生是复杂的,不能理解为一个单个的原子或基团就使一种化合物成为抗性的,而实际抗性形成能力是十分复杂的。
和增强抗性的基团不同,也有一些基团具有促进生物降解作用,一般认为如下的情况可以促进生物降解。①存在酶催化水解的潜在位点,如酯、酰胺;②化合物能通过导入氧形成羟基、醛基和羧酸;③存在未取代(unsubstituted)的线性烷链(特别是≥4C)和苯环。其中第二种情况最为重要,因为对许多化合物(如烃)来说生物降解中的第一步是酶催化把氧插入到结构中,并且常常是降解速率的限制步骤。
附加在化合物基本结构如苯环上的取代基的数量和位置对生物降解具有一定程度的影响。取代的程度和生物降解性有确切的联系和指示价值,如间位取代苯比邻位、对位取代难降解,2,4,5-T比2,4-D难降解得多,但一些化合物被取代以后反而更易降解。但总体上难以形成可用于预测的一般性规律。
化合物功能基团对生物降解的影响也被用空间效应和电子效应来解释。空间效应是化合物降解过程中降解酶与底物相接触反应位点被分枝或功能基团堵塞后降解反应速率降低的现象。8碳的直链烷烃辛烷的两端被氧化成醇或过氧化物,然后被进一步降解,而有甲基支链的同样8碳烷烃就会抑制开始于两端的降解(两种结构如图8-45)。使化合物难以降解。
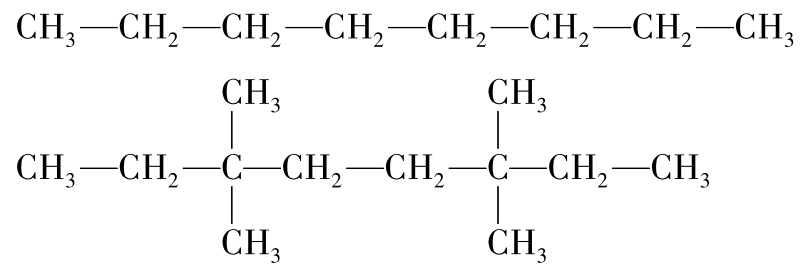
图8-45 两种空间结构不同的烷烃结构
此外支链或异构功能基团还能影响底物透过细胞膜的运输,特别是当这种运输是连接到酶上的时候,空间效应在功能基团的体积增大通常会得到加强。电子效应是功能基团通过反应位点电子密度的改变而影响生物降解。功能基团可以通过吸电子(如Cl)和供电子(如CH3)来改变反应位点的电子层密度。通常增大反应位点电子密度的功能基团能提高生物降解速率,而减少反应位点电子密度的功能基团则降低生物降解速率。有人比较研究了一系列邻位取代苯酚的电负性和它们降解速率关系,通过对五个不同功能基团的测试发现在取代基电负性增加时,生物降解速率下降(图8-46)。
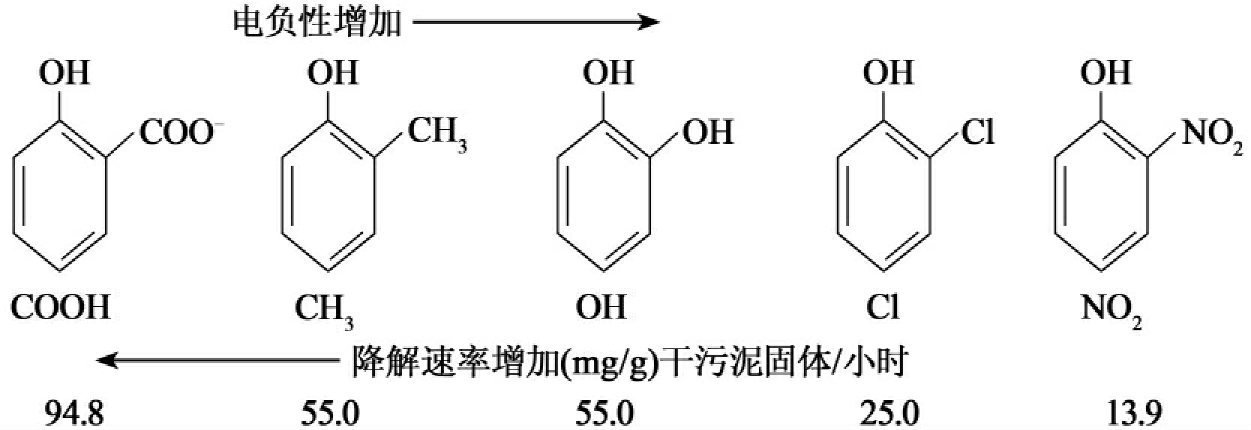
图8-46 不同的邻位取代苯酚及其降解速率
有关学者在大量试验研究的基础上提出了能较快降解的化合物的结构模式。
①分子量少于74的所有化合物。
②不带环的所有单元酸。
③所有由C、H、N和O原子组成,有1,3或更多的C—O键,但不带季碳的化合物。
④所有由C、H、N和O原子组成,有至少一个C—O键,分子量大于103U,但没有季碳的化合物。
⑤分子量大于103U,有1,3或更多C—O键,2或3环的化合物。
⑥分子量大于103U,由C、H、N和O组成,3环或少于3环,但没有季碳,也没有芳香氨基的化合物。
⑦分子量大于103U,由C、H、N和O组成,有2个或3个环,但没有季碳。
二、预测生物降解性定量结构生物降解性关系模型的构建
一种化合物的生物活性取决于分子结构及其他生物和环境因素,但分子结构是决定性的,起主要作用的因素,生物活性包括生物降解性、生物毒性及其他性质。分子结构与生物活性的关系被称为QSARs(quantitative structure activity relationships),1994年QSARs首先用于生物降解,用于生物降解方面的QSARs(QSARs for biodegradation)就成为QSBRs(quan-titative structure-biodegradability relationships),也称为SBRs(structure-biodegradability relationships)。一种化合物的生物降解受多种因素的影响,主要是化合物的自身结构、化合物理化性质、降解微生物以及所提供的环境条件。如果能提供充分驯化、适应的微生物及合适的环境条件,则化合物的降解在很大程度上就取决于化合物自身,化合物的自身结构及物理、化学特征是影响生物降解的根本因素,化合物分子结构的信息可以反映在生物降解上。微生物及环境条件是外因影响降解的外部条件。
基于分子结构对生物降解的重要性和结构与生物降解的这种定量关系(QSBRs),科学研究人员就逐步发展出以分子结构中基团为基础的预测化合物降解性的预测用的降解数学模型,利用这些模型可从分子构成特征出发来预测化合物的生物降解性,从而可以进行风险、归宿评价,并为进一步设计环境安全的化合物服务。
1.模型建立的基础
假设生物降解性是组成化合物分子的一个或更多基团(片段)所贡献的功能,并且每种基团对各种化合物来说是不变的(即设定同一片段在不同化合物中起同样作用,忽略不计片段之间的相互作用)。理想的情况是这种模式中的每个片段对降解活性都有清楚的机理关系。尽管孤立地看待结构与生物降解性之间的关系是不科学的,但通过一定的方法可以消除这些偏差,而且也可以把两个基团或更多基团及它们之间的相互作用作为一类新的基团加以考虑。这样我们就可以发展出一种模型,在这种模型中,生物降解性用一系列的合适大小的可计算值表示,同时以某种方式把生物降解性和结构片段结合起来产生出合理的综合模型。我们以化合物结构为基础的模型仍然存在很多局限性,但有这些模型进行预测要比没有好得多。建立模型也可以认为是对新化合物风险管理和规避风险的一种方法。
2.建立模型的程序
构建有机化合物定量结构生物降解关系(QSBR)模型的程序一般经过6个主要步骤。
(1)选择系列化合物
选择一系列不同类型和不同取代基位置的化合物,化合物取代基常数的值应具有广谱范围的性质,即在选定的取代基参数之间其变化相关性应最小。
(2)分子结构及生物降解试验
选择合适的结构参数研究较为独立的分子结构描述符,确定获取生物降解性速率的测试方法。
(3)分子结构参数的转换和生物降解性速率的获得
分子结构参数转换成数字描述。生物降解速率的获得必须在相同条件下进行,这包括生物降解的终点、方法及所提供的环境条件,获取尽可能多的生物降解性能数据。
(4)建模
选择合适的方法建立生物降解性和结构参数的定量关系模型。采用诸如回归分析等数学统计方法,去除生物降解性影响小的参数,保留重要参数。
(5)模型检验
以大量的化合物的实例结果检验模型,并不断校正模型优化模型,给出模型的适用约束和误差范围。
(6)实际应用
预测新的化合物的生物降解性,评价其归宿、行为,并为合成新化合物提供支持。
3.建立模型的方法
所有能影响生物降解性的结构及物理、化学特征都能作为构成QSBR的参数,现在许多研究QSBR的学者都从不同的角度把影响生物降解因素综合到生物降解中,构成QSBR来预测化合物的生物降解性。
何菲等把目前较为常见的QSBR建模方法概括为:线性自由能相关法(LFER,Hansch分析法)、基团贡献法(free-wilson法)、分子连接性指数法(MCI法)、专家系统(expert system)、人工神经网络法(artificial neural networks,ANN)、比较分子力场分析(comparative molecular field analysis,COMFA)等。
线性自由能相关法(linear free-energy relationship,LFER):这是QSBR研究最为常用的方法,由于Hansch研究得最为深入,故而又称其为Hansch分析法。其理论基础是基质分子结构的微小改变将导致限速步骤活化能的线性改变,进而影响降解速率的改变,用数学式表达为:
lgK=A1X1+A2X2+…+AnXn+C
式中,K为生物降解速率常数,A1-An为系数,X1-Xn为有机物分子结构描述符,C为常数。应用这种方法许多学者得到一大批预测模型。
基团贡献法:这种方法是在对化合物亚结构信息和生物活性相关研究的基础上建立的。该法将各种化合物分子按其结构分解为几个官能团或片段,假定每个官能团或片段对化合物的生物降解都有特殊的贡献。生物降解速率常数K可用贡献函数X表达,对于化合物的每一个基团或片段都可以使下式成立:
ln(K)=f(a1,a2,…,aj)
用泰勒级数(Talor Series)将上式展开,若忽略二阶以上的部分,即可获得生物降解速率常数K的一级线性模型,表达为:
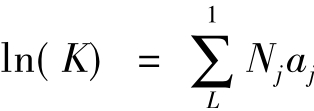
式中,Nj为化合物中第j类基团的数目j为第类基团的贡献值,L为化合物中基团的总数。
对于每一种化合物,都可建立一个这样的线性方程,应用最小二乘法可以解出K值。这种模型只用了一级近似,但如果基团之间的相互作用很重要,就不能使用这样的模型,可考虑使用二阶或更高阶的方程处理。
分子连接性指数法:分子连接性指数(MCI)方法是目前最常用的建模方法之一。以MCI指数建立同生物降解性间的线性相关方程,就是MCI-QSBR法,这种方法的优点是完全从化合物的分子信息着手,而不必考虑微生物降解的代谢途径与限速反应,因而可能使QSBR不仅仅适用于同系或同族化合物,而且也适应于许多其他化合物。有学者以该法对29种不同类的芳香化合物建立了非驯化的生物降解模型,得到如下方程:
lgK=0.85(0x-0xv)-6.51(5xcv)+0.72G-0.89A+0.60
n=29,r=0.93
式中,G与A分别为基质与脂肪侧链的指示变量。
专家系统:专家系统对化合物的生物降解性的认定主要依赖于少数技术专家的职业判断。降解数据不是直接的实验研究结果,而是专家的推理及经验的判断。由于缺乏实验数据,对许多化合物来说不确定性仍然很大,这使生物降解途径与速率并不完全明确。但专家的认识从本质上来说是来源于基团对生物降解性的贡献的认识,专家预测生物降解性,常以基团贡献法为基础,特别注意促进降解性和阻滞降解性的结构片段。同时可能结合几类参数,例如理化参数、分子连接指数等。因此专家系统实际上是一种结构生物降解性的人工智能判断。
人工神经网络法:人工神经网络是模拟人脑结构的一种大规模的并行连接机制系统,具有自适应建模学习和自动建模功能。特别对线性问题有良好的拟合预测能力。
比较分子力场分析:比较分子力场分析是最重要的3D-QSARL三维定量构效关系,即基于分子的三维结构对其性质或活性进行预测方法之一。这种方法将一组具有相同性质(降解活性)的分子按照其相同的几何作用点,在三维空间进行叠加,计算这一组分子叠加的立体场和静电场,用某种探针原子对这些场进行作用,然后用偏最小二乘(PLS)及交叉验证得到预期模型。即通过比较活性化合物与非活性化合物的有关分子结构信息,可以筛选并确定对分子生物活性起关键作用的化合物电子结构或立体结构特征,进而推测化合物-受体作用机制,建立化合物生物降解模型。
4.表征结构及理化特征的描述符
QSBR模型实际上是把有机化合物的结构性质(或理化性质)与其生物降解性之间的关系用数学模型加以解析、表达的过程。有机化合物的结构用化学结构描述符予以表征,目前常用的主要有:
①理化性质描述符:包括分子量(MW)、正辛醇/水分配系数(Kow)、酸解常数(pKa)、碱解常数(KOH)、分子表面积(TSA)、高压液相色谱保留时间(RT)、疏水常数(π)、溶解度和分子连接指数。
②电子效应描述符(电子参数):包括电子效应参数和量化参数。
③空间效应描述符(空间参数):空间效应反映的是取代基的位阻效应,这一效应可改变酶反应中心和化合物的接触。空间效应的参数主要有分子量、范德华半径(Yw)、Taft常数(Es,Es越负,其对氢原子的空间需求越高,即位阻越高)等。
5.生物降解性预测
生物降解性预测是利用模型把化合物分子结构的信息转化成生物降解速率数据的过程。预测要提供二种类型的生物降解资料,第一种资料是化合物完全降解还是持久存在,对于持久性化合物要回答其在环境中的积累潜力。从模型得到的数据还要结合环境的条件,把理论上的数据转化成在实际的真实世界中的可能行为。
6.模型的发展
应用人工智能计算机程序构建有机化合物的结构-生物降解关系的数学模型,是构建更好模型的发展方向。MULTICASE公司开发出建立在化合物分子结构基础上的预测生物降解产物、代谢产物的潜在毒性和生物降解性的META-CASETOX系统。这个系统含有能预测化合物生物转化形成代谢产物的META计算机程序。程序实际上是一个带有转化规律词典、multiCASE人工智能支持的专家系统。系统能自动识别特定化合物的亚结构片段(substructural fragment)和替代片段以及片段的生物活性(降解活性)。当一种新的化合物被提交到程序时,所有可能亚结构片段被识别,并与储存在词典中的识别片段比较。当一种相似物被认定时,程序对母体化合物执行这种规则产生结果代谢物,每种代谢物能被进一步分析,这样产生了代谢物树。对适用不同规则的结构,可根据已知的实验资料预先设定优先的转化规律。程序数据库中好氧降解数据由385种分子结构组成,涵盖多样的降解原理和化学模型。每种化合物活性相关成生物活性值,这种值可以是定性的,即具有活性或不具有活性(是可生物降解性或不被生物降解性);也可以是定量的,如降解速率、反应速率常数、理论需氧量百分数(%THOD)、生物降解半衰期(T50)等。亚结构片段和可经程序计算出来的化合物的亲脂性、水溶性、空间指数、量子力学指数作为变量,再经过线性回归分析就得到QSBR模型。
Activity=a+∑bi(niFi)+clgP+dlg2P+eMwt+fws+gGi+h(QM)
式中:a,b,…,h为回归系数,ni是一种片段i出现在一种化合物中的次数,如果片段i存在;Fi等于1,否则为0,lgP是n-辛醇和水之间的分配系数的一般对数。Mwt是分子量,ws是水溶解度,lgP和ws作为化合物的迁移特征包括在模型中。Gi是拓扑学graph指数。QM代表另外的量子力学参数,包括HOMO和LuMo系数。
导出可靠和更好的可应用QSBR模型,要特别重视化合物在野外经历的生化转化途径,尤其注意可能形成的截止式产物(dead-end products)、持久性中间产物及其毒性潜力。这些相关化合物和它的代谢物的毒性应作为QSBR模型的一个描述符。
7.模型的局限性
应用模型来预测化合物生物降解性从本质上说是一个把有限的实验认知、试验数据、所得到的结论外推到一个有无穷变化的真实环境的过程。挑战是可否得到代表真实的数据和如何把可利用数据外推到相关的真实世界。这就存在一种不可预知性和不确定性,这就是模型的局限性。
①理论认识上的局限性:我们对生物降解的机理、过程以及影响因素的理解认识远远不够,存在着很大的局限性。
②数学模型的不完整性:化合物在环境中的生物降解过程是一个非常复杂的过程,除微生物的作用外,其他的环境因素都对生物降解产生重要影响。实验室得到的试验结果外推到实际的环境是一个更复杂的过程,而目前的模型不能反映这个复杂过程,所以模型缺乏完整性也是局限之一。
③构建模型试验数据的不真实性:实验室和野外模拟实验的数据是建立模型的基础,但实验条件下的数据相对真实环境来说往往是不真实的,因而在建立模型外推过程中存在不确定性。首先至今我们未对影响降解的各种因素进行彻底的研究和完全的了解。此外标准的实验室实验和模拟试验并不类似于野外状况,反映真实环境。例如一般生物降解试验都用野外环境所不存在的高浓度的试验基质和高浓度的培养物。因此试验应尽可能接近野外基质存在的环境条件。例如高疏水性化合物的降解试验在厌氧条件下进行,而亲水非挥发性基质的降解试验在好氧水环境中展开。这将不仅有助于把实验结果外推到真实环境,也有助于发展QSBR能力去预测化合物在真实环境中的行为。再就是建模来源于不同的试验研究目标,试验大多零散,不系统,因此数据的数量不足和缺乏可比性。
三、定量结构生物降解模型的应用
以化合物结构参数为基础的生物降解模型主要有二个方面的应用,一是对化合物进行筛选,列出顺序,以便作出评价,二是对设计期化合物作出筛选。
1)把化合物的分子基团代入模型可以算出一个与生物降解性相关的数值。这个数值可以指示化合物的可降解性,从而可对大量化合物的生物降解性排序,为化合物的风险管理和规避风险提供技术支持。
2)设计可生物降解化合物。基于下述理由可生物降解化合物的设计是极为重要的。
①从源头上降低污染的思维方式是设计安全化合物的直接推动思想。②生物降解是土壤、水体和污水处理中大部分有机化合物主要的失去机理。③生物降解性被作为产品设计的一个参数,并与毒性、化合物的功能被放在一起考虑。通过综合(掺合)更大的生物降解性到非毒性产品的分子结构中,环境损害的风险被减少。实际上,生物降解性已经成为一种重要的设计考虑,这种考虑对日常消费品(如日常的化工产品)已引起注意。而在一些工业产品的设计和生产中尚未得到重视。
下面的例子特别说明结构与生物降解性的关系,并说明BIODEG模型(一种QSBR模型)可被用于化合物底物生物降解性预测。
线性烷基苯磺酸盐的发展使用是说明分子结构与生物降解性相互关系的最显著的例证,其充分说明分子工程学对促进生物降解的重要作用,并说明这样的化合物可被环境接受。20世纪40年代人造的烷基苯磺酸盐(alkybenzene sulfonate,ABS)替代了肥皂。开始烷基链是从煤油中来,但很快就被丙烯四聚物(propylene tetramer)代替。四丙烯烷基苯磺酸盐(tetrapropylene alkylbenzene sulfonate,TPBS)是一种更有效和经济的产品,其是通过one-step friedel-crofts process得到的。这种高度分支产品的环境问题很快就表现出来,因为它们不能在城市的污水处理系统中被完全降解,相关研究证明TPBS仅能被降解大约50%,结果在活性污泥污水处理槽和受纳水体中产生过量泡沫,而且还以较高的浓度(高达2mg/L)存在于受纳水体中。此外泡沫的存在影响污水处理的有效性,增加处理成本。
新的对环境友好的表活剂LAS在20世纪60年代又取代了TPBS。LAS的线性烷烃是用分子筛(molecular sieves)从石油中得到的。
LAS表面活性剂在污水处理厂中几乎可被完全降解。与LAS相比,TPBS较低的生物降解性是由于它的高的分支烷基,而LAS的较大的促进生物降解是由于缺乏这种分支。线性LAS促进生物降解易于从BIODEG模型中预测,LAS含有促进生物降解的片段(这不存在于TPBS中),这就是带有≥4碳的线性末端烷基。
从化合物的分子结构与生物降解性的大量研究可以看到设计更易生物降解化合物的方法是综合促降解的正性因素(如酯键和羟基)、排除抗降解的负面因素(如卤素、叔碳、硝基)。
一种产品的生物降解性和经济性有明显的密切联系。化合物的产品设计实际上是一个多学科的过程。化学品的制造者、拥有者和使用者必须更艰难面对它们的经济问题部分,但这种平衡过程可能受损害。新增的经济性或功能会造成一种新的耗费,而这些经济性或功能是先前所不需要的。这种改变被看做内化(internalizing)环境成本过程的一部分。把完整的环境考虑的愿望放到商业决定中(选择),设计产品使其环境影响达到最低现在已不成为问题,并已在CMAK'S Responsible care program中确认。
我们必须使所有的消费品和工业品都应反映扩大到可实践的安全设计原理中。有4种理由说明我们为什么要提出这种目标:
①我们不能预先就知道释放的化合物的可能毒性效应,这不但对研究较少的新化合物如此,有些得到很好研究的化合物也是可能的,壬基酚羟乙基盐争论就说明这一点。
②一种化合物会释放到环境中,如果产品是成功的则会明显增加释放量。
③随着时间的推移,一种产品的市场份额会不断增大,例如具有非常有限市场生态位的表面活性剂可以在更大范围找到它们的用途(在纺织过程、造纸或其他工业部门中应用)。
④全球市场的开放进一步扩大,全球环境相联系,一个国家生产的产品可以输出到其他国家,输出到环境控制较不苛刻的环境,造成大范围迁移的可能性,环球损害可以不被限于释放的地点。
思 考 题
1.试述研究微生物对环境有机污染物降解转化的重要意义。
2.生物降解机理是什么?
3.生物降解的基本条件是什么?
4.影响生物降解的环境因素有哪些?
5.怎样认识和理解微生物降解有机污染物的巨大潜力?
6.举例说明降解质粒在难降解有机污染物生物降解中的重要作用。
7.研究生物降解遗传进化在理论和实践上有何重要意义?
8.生物降解遗传进化的机理是什么?
9.有机污染物降解过程中的主要降解反应包括哪些?
10.试述有机污染物生物降解速率的不同表示方式及其意义。
11.试述环境污染物在环境中的行为以及影响因素。
12.试述脱卤酶催化脱卤的主要方式。
13.降解菌及其降解基因的指示系统主要有哪些?
14.降解酶的工程化改造方法有哪些?
15.以TCE为例说明共代谢降解过程。
16.共代谢降解的主要特点是什么?
17.共代谢过程的应用是什么?
18.比较不同脂肪烃化合物的生物降解性并从中找出其规律性。
19.生物降解性测定的主要测定方法有哪些?
20.生物降解性测试中受试化合物浓度及注入方式如何确定?
21.试述生物降解性研究的完整过程。
22.说明抗降解性和促进降解的结构特征。
23.怎样认识预测生物降解性模型的局限性?