自然地理学
-
1.1第一章 地球演化
-
1.1.1第一节 地球的形成与演化
-
1.1.1.1一、相对地质年代的确定
-
1.1.1.2二、绝对地质年代的确定
-
1.1.1.3三、地质年代表
-
1.1.1.4四、地球的演化历史
-
1.1.2第二节 地质构造
-
1.1.2.1一、内力作用及其表现形式
-
1.1.2.2二、地质构造形式
-
1.2第二章 岩石圈系统
-
1.2.1第一节 大陆岩石圈
-
1.2.1.1一、地壳盖层
-
1.2.1.2二、上地壳刚硬层
-
1.2.1.3三、中地壳塑性层
-
1.2.1.4四、下地壳刚硬-软弱复合层
-
1.2.1.5五、莫霍面过渡带
-
1.2.1.6六、地幔盖层
-
1.2.2第二节 大洋岩石圈
-
1.2.2.1一、未固结沉积层(层1)
-
1.2.2.2二、火山岩层(层2)
-
1.2.2.3三、玄武岩层(层3)
-
1.2.2.4四、壳幔过渡层
-
1.2.2.5五、浅地幔刚硬层
-
1.2.3第三节 岩石圈的基本构造单元
-
1.2.4第四节 板块构造
-
1.2.4.1一、板块概念与板块划分
-
1.2.4.2二、板块边界
-
1.2.4.3三、板块运动的全球图谱及运动速率
-
1.2.4.4四、板块运动的观测方法
-
1.3第三章 地貌过程
-
1.3.1第一节 经典地貌发育理论
-
1.3.2第二节 地貌发育系统
-
1.3.2.1一、地貌发育的内动力
-
1.3.2.2二、地貌发育的外动力
-
1.3.2.3三、内外力相互作用
-
1.3.3第三节 地貌形体类型
-
1.3.3.1一、地貌形体
-
1.3.3.2二、地貌类型
-
1.4第四章 地貌系统
-
1.4.1第一节 岩石地貌系统
-
1.4.1.1一、砂质岩石地貌
-
1.4.1.2二、喷出岩地貌
-
1.4.1.3三、花岗岩地貌
-
1.4.1.4四、岩溶地貌
-
1.4.1.5五、黄土地貌
-
1.4.1.6六、生物岩地貌
-
1.4.2第二节 动力地貌系统
-
1.4.2.1一、构造地貌
-
1.4.2.2二、坡地重力地貌
-
1.4.2.3三、流水地貌
-
1.4.2.4四、冰川地貌
-
1.4.2.5五、冻土地貌
-
1.4.2.6六、风沙地貌
-
1.4.2.7七、海岸地貌
-
1.5第五章 地球大气
-
1.5.1第一节 大气成分与结构
-
1.5.1.1一、大气成分
-
1.5.1.2二、大气圈的垂直结构
-
1.5.2第二节 大气能量
-
1.5.2.1一、大气辐射平衡
-
1.5.2.2二、大气热力均衡
-
1.5.3第三节 大气运动
-
1.5.3.1一、大气运动的驱动力
-
1.5.3.2二、大气环流和风系
-
1.5.3.3三、气旋和反气旋
-
1.5.4第四节 大气降水
-
1.5.4.1一、大气湿度
-
1.5.4.2二、水汽凝结
-
1.5.4.3三、大气降水
-
1.6第六章 气候与环境
-
1.6.1第一节 气候形成
-
1.6.1.1一、气候形成的太阳辐射因素
-
1.6.1.2二、气候形成的大气环流因素
-
1.6.1.3三、气候形成的地表环境因素
-
1.6.2第二节 气候分异
-
1.6.2.1一、气温分异
-
1.6.2.2二、湿度和降水分异
-
1.6.2.3三、气候分异
-
1.6.3第三节 气候类型
-
1.6.3.1一、柯本气候分类
-
1.6.3.2二、斯查勒气候分类
-
1.6.3.3三、世界气候类型
-
1.7第七章 水循环与水分运动
-
1.7.1第一节 地球上水的分布
-
1.7.2第二节 水循环与水量平衡
-
1.7.2.1一、水循环
-
1.7.2.2二、水量平衡
-
1.7.3第三节 水分运动和输送
-
1.7.3.1一、海水的运动和输送
-
1.7.3.2二、径流的流动与输送
-
1.8第八章 地球水环境系统
-
1.8.1第一节 海洋水环境
-
1.8.1.1一、海水的性质
-
1.8.1.2二、海洋的组成
-
1.8.1.3三、海洋形态结构
-
1.8.1.4四、海洋对地理环境的影响
-
1.8.1.5五、海平面变化
-
1.8.1.6六、海洋荒漠化问题
-
1.8.2第二节 陆地地表水环境
-
1.8.2.1一、河流
-
1.8.2.2二、湖泊
-
1.8.2.3三、沼泽
-
1.8.2.4四、冰川
-
1.8.3第三节 地下水环境
-
1.8.3.1一、地下水的概念
-
1.8.3.2二、地下水的类型
-
1.8.3.3三、地下水的动态与平衡
-
1.8.3.4四、地下水污染
-
1.9第九章 生物圈系统
-
1.9.1第一节 生物圈与生物多样性
-
1.9.1.1一、生物圈
-
1.9.1.2二、生物多样性
-
1.9.1.3三、生物与环境
-
1.9.2第二节 生物群落
-
1.9.2.1一、群落
-
1.9.2.2二、群落结构
-
1.9.2.3三、群落演替
-
1.9.3第三节 生态系统的组成与结构
-
1.9.3.1一、生态系统的概念
-
1.9.3.2二、生态系统的组成成分
-
1.9.3.3三、生态系统的结构
-
1.9.4第四节 生态系统功能
-
1.9.4.1一、生物生产
-
1.9.4.2二、能量流动
-
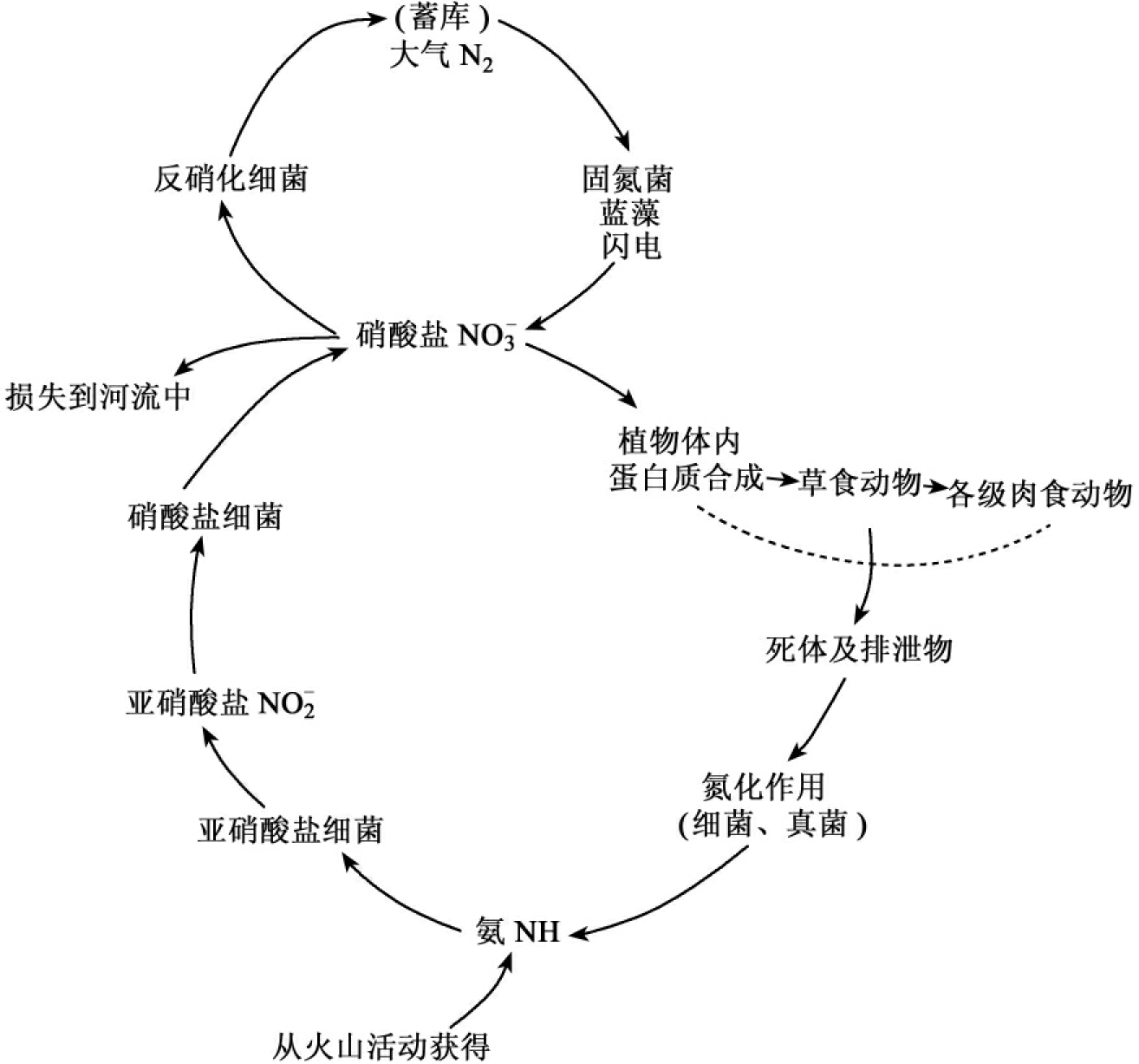
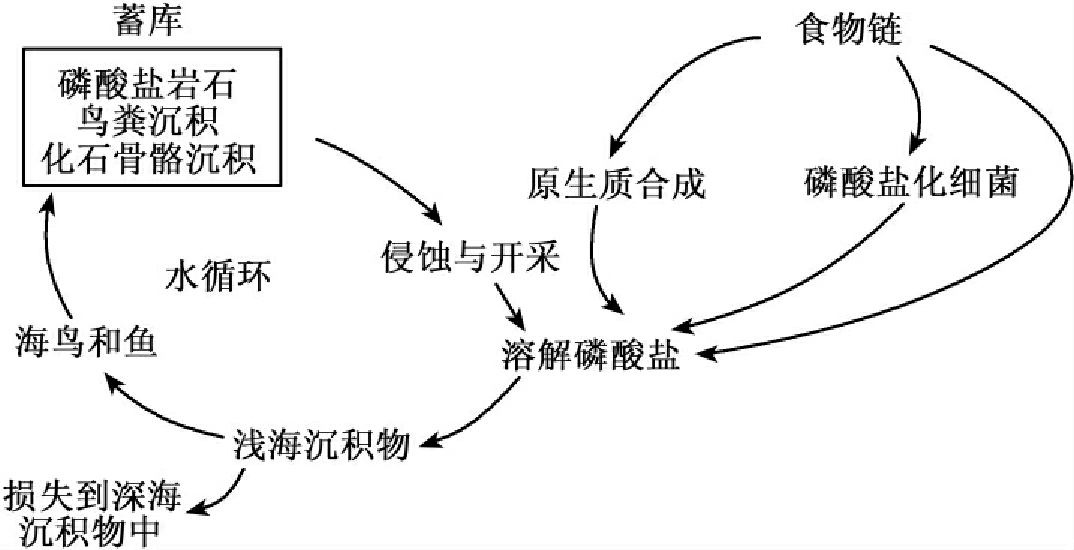
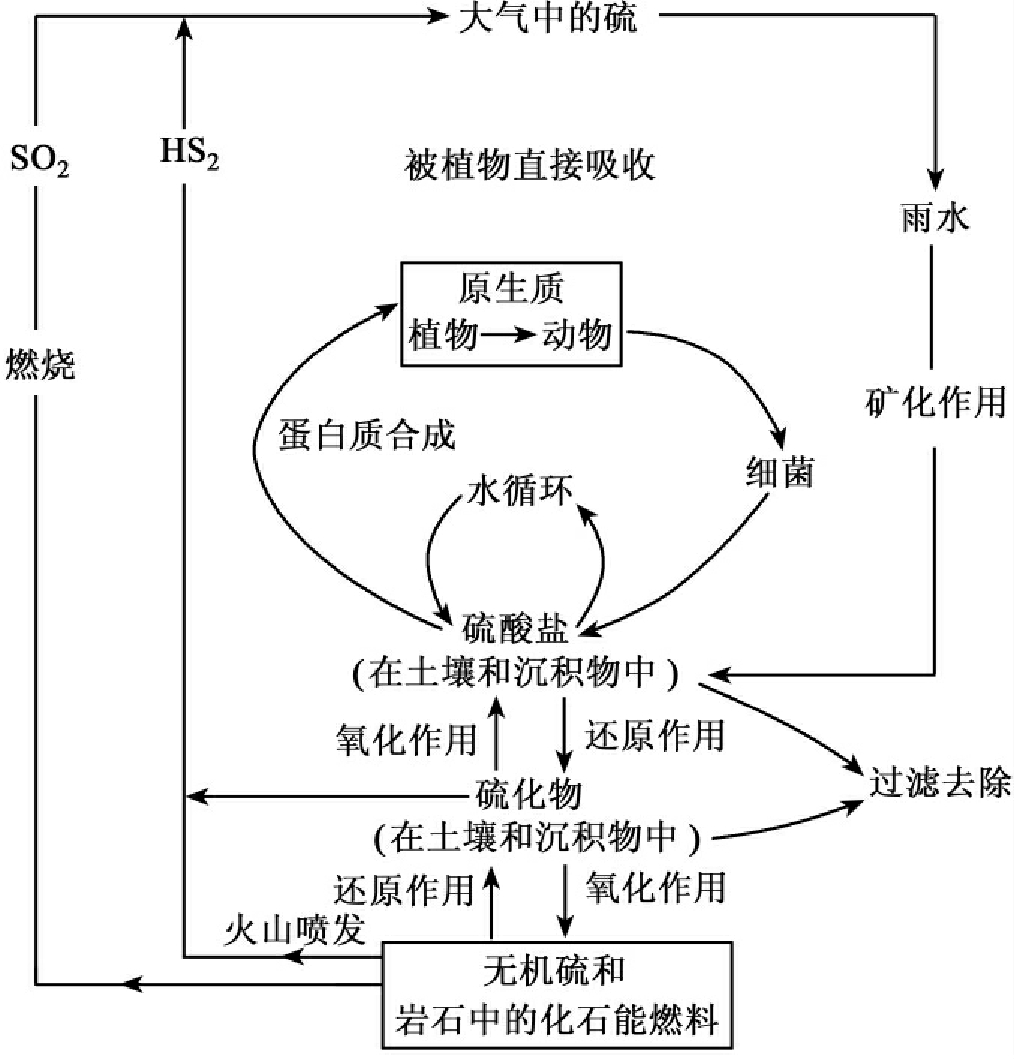
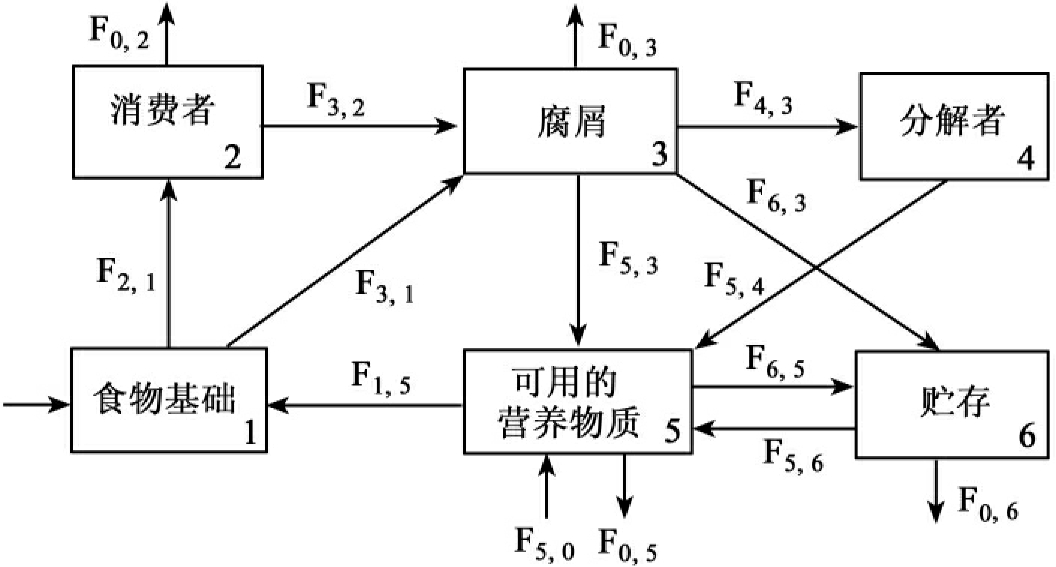
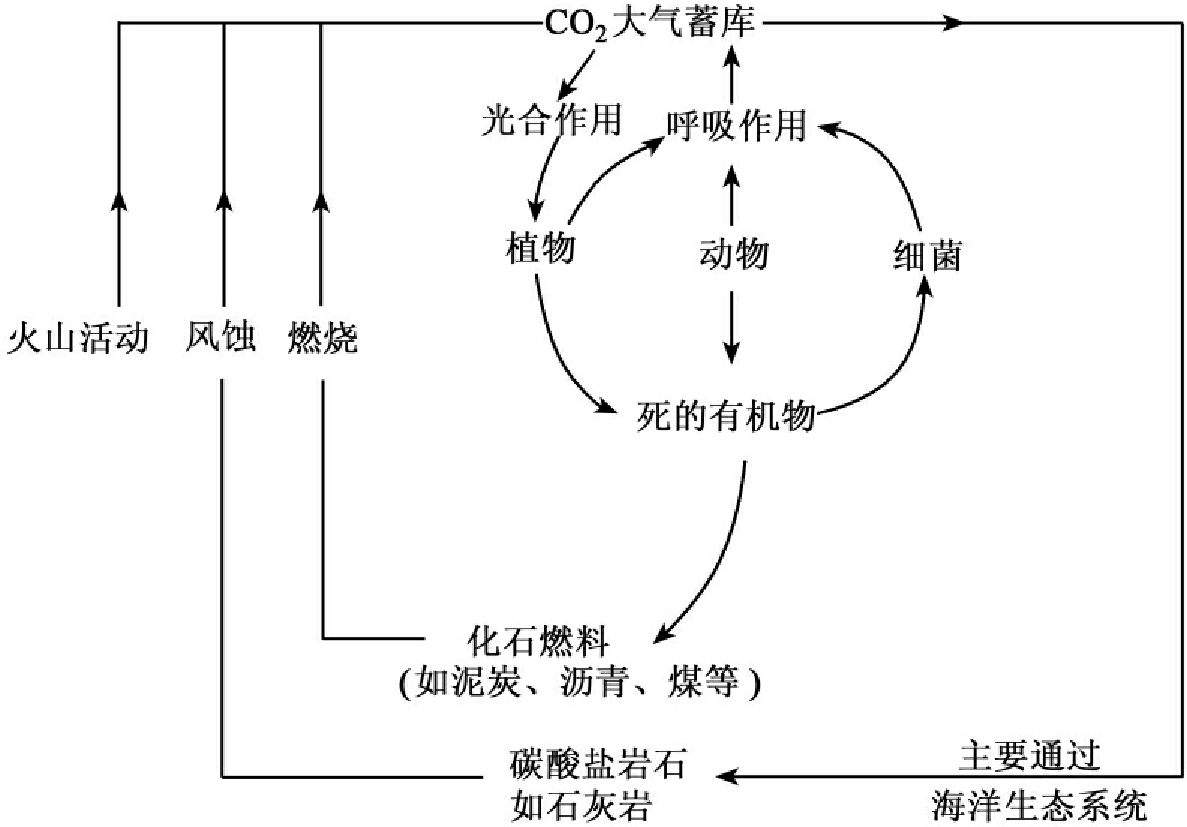
1.9.4.3三、物质循环
-
1.9.4.4四、信息传递
-
1.9.5第五节 生态平衡
-
1.9.5.1一、生态系统的反馈调节
-
1.9.5.2二、生态平衡
-
1.9.6第六节 生态修复
-
1.10第十章 地球上的生态系统
-
1.10.1第一节 森林生态系统
-
1.10.1.1一、热带雨林生态系统
-
1.10.1.2二、常绿阔叶林生态系统
-
1.10.1.3三、落叶阔叶林生态系统
-
1.10.1.4四、北方针叶林生态系统
-
1.10.2第二节 草原生态系统
-
1.10.3第三节 荒漠与苔原生态系统
-
1.10.3.1一、荒漠生态系统
-
1.10.3.2二、苔原生态系统
-
1.10.4第四节 湿地生态系统
-
1.10.4.1一、湿地的特点
-
1.10.4.2二、湿地的功能
-
1.10.4.3三、湿地的价值
-
1.10.5第五节 河流、湖泊生态系统
-
1.10.5.1一、河流生态系统
-
1.10.5.2二、湖泊生态系统
-
1.10.6第六节 海洋生态系统
-
1.10.6.1一、浅海带生态系统
-
1.10.6.2二、外海带生态系统
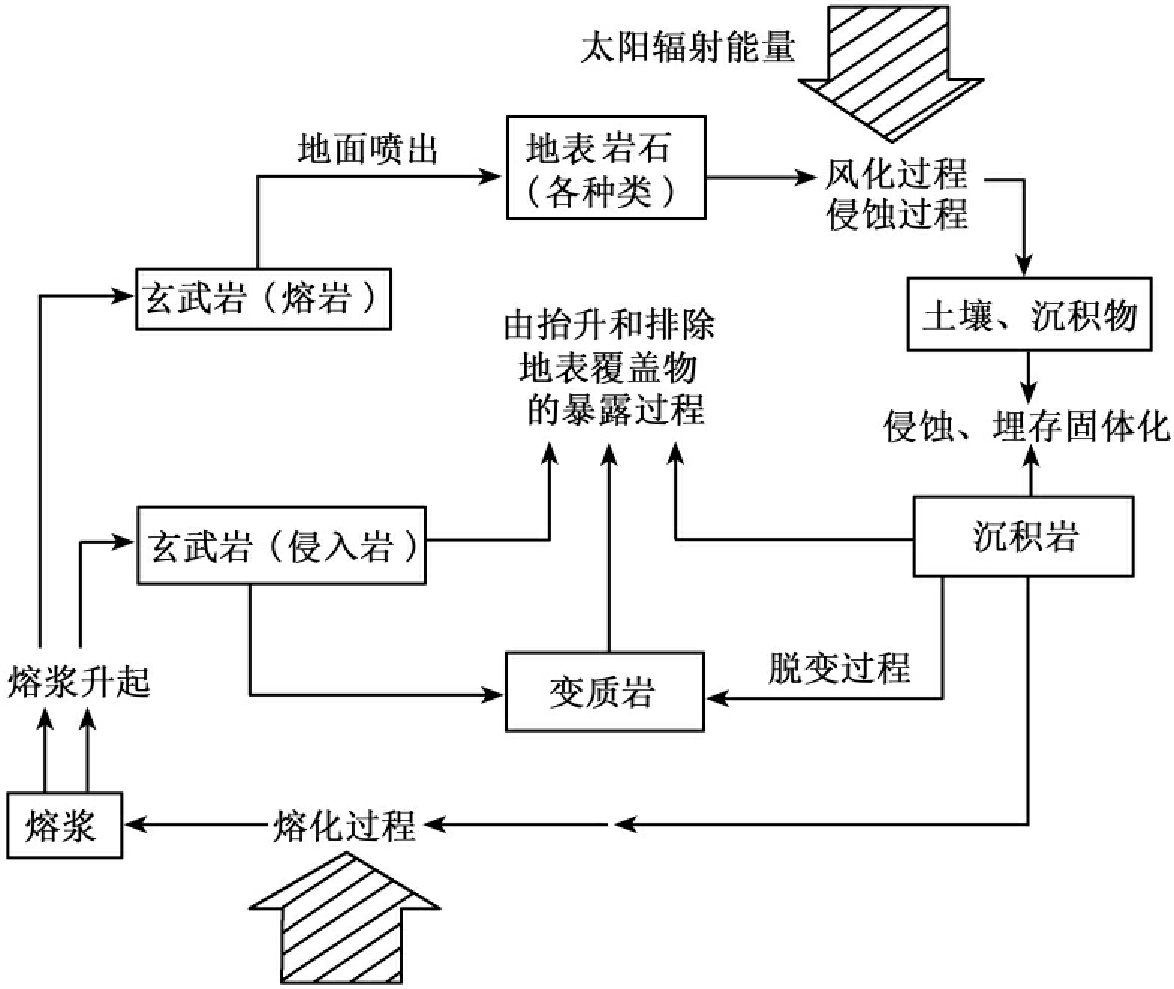
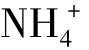 ,可为植物所直接利用。
,可为植物所直接利用。