13 以生理为基础的人格维度
我们已经介绍了人格研究中使用的一些基本生理测量,下面将转到某些令人格心理学家感兴趣或引起他们注意的理论。首先从最广为人知的人格生理学理论开始,该理论提出了一种生理学的解释:为什么有的人内向,而有的人外向?
外向性—内向性
在你认识的人中,一些人可能符合下面的描述:健谈、友好、喜欢交新朋友、去新地方、精力充沛、有时冲动和冒险、容易厌烦、不喜欢老套和单调。这样的人在完成外向—内向人格量表时,在外向性上得分高。表2列出了流行的艾森克人格量表中有关测试内向和外向的题目。
也许你还认识一些人,他们可能恰恰相反:安静、退缩,在人多的时候喜欢独处或与少数几个朋友在一起,做事有规律有计划,喜欢常规而不是意外。这样的人在完成外向—内向人格量表时,在内向性上得分高。如果你纳闷为什么内向者和外向者如此不同,生理学取向的人格心理学家给出了一个很令人感兴趣的解释:艾森克的理论。
表2 艾森克人格问卷中外向性的测试题目

计分方法:将带有“(反)”的题目进行反向计分。然后,统计回答“是”的数量。大学生在这个问卷上的平均得分是11分。
1967年,艾森克在《人格的生物学基础》一书中提出了生理取向的人格理论的一种经典范例。艾森克认为,内向者的大脑上行网状激活系统(ARAS)的活动水平比外向者高。ARAS位于脑干,控制着整个大脑皮质的唤醒水平。在20世纪60年代,ARAS被认为是神经刺激进入大脑皮质的门户。如果这个门户的开放程度低,那么大脑皮质的基线唤醒水平就较低;如果其开放程度高,那么大脑皮质的基线唤醒水平就高。照此理论解释,内向者的大脑皮质有较高的基线唤醒水平,因为他们的ARAS允许太多的刺激进入。内向者之所以表现出内向行为(安静、喜欢低刺激的环境,如图书馆),是因为他们需要使已经较高的大脑皮质唤醒水平得到控制。相反,外向者的外向行为是为了提高大脑皮质的唤醒水平。

你是一个健谈者吗?你喜欢和人交往吗?你喜欢整天忙碌和充满刺激吗?对这些问题说“不”,说明你是一个内向者。
艾森克还将赫布的“最佳唤醒水平”的观点整合进了自己的理论。赫布认为,最佳唤醒水平是指正好适于某项任务的唤醒水平。例如,试想你在一个低唤醒状态下(磕睡、疲惫)参加期末考试。此时,低唤醒的状态同过度唤醒(如焦虑和激动)的状态一样糟糕。参加考试需要一个最佳的唤醒水平:注意力集中、适度紧张、专心,但不要达到焦虑状态。

你喜欢给朋友讲笑话或有趣的故事吗?你喜欢扎人堆吗?你能使一个聚会进行下去吗?对这些问题说“是”,说明你是一个外向者。有趣的是,艾森克的内向—外向理论不是基于对与人相处的需要,而是基于对唤醒和刺激的需要。
如果内向者的基线唤醒水平(即放松状态下的唤醒水平)高于外向者,那么内向者高于最佳唤醒水平的状态要多于外向者。根据艾森克的理论,总是处于过度唤醒状态使内向者采取更多的控制或压抑策略,他们回避积极的社会交往,因为这将增强已经过高的刺激水平。另一方面,外向者需要提高唤醒水平,所以总是寻求刺激性的活动,做出一些不受约束的行为。典型的内向者安静、退缩,典型的外向者好动、忙碌。他们这样做是为了使其唤醒水平降低(内向者)或升高(外向者),以维持各自的最佳唤醒水平。
在艾森克的理论发表后的数十年里,许多研究对此进行了验证。如果内向者的皮质唤醒水平真的比外向者高,那么,内向者应该表现出脑皮质活动反应的提高,如脑电图的增强;以及交感神经系统活动的增加,如皮电反应。为了检验这个经典假设,研究者通过在不同的刺激条件下得到的生理指标,对内向者和外向者进行了比较。在无刺激或弱刺激条件下,内向者和外向者之间没有差异或差异很小。但是在中等刺激条件下,内向者的神经系统的反应比外向者更大或更快,这与依据艾森克理论所做的预测一样。
内向者和外向者在基线唤醒水平上没有不同,而在中等刺激条件下存在差异,这个事实使艾森克修改了他的唤醒理论。1967年,艾森克首次提出他的理论时,并没有对静息水平(或基线水平)、唤醒水平与唤醒反应进行区分。现在有大量证据表明,内向者和外向者的真正不同在于唤醒能力或者唤醒反应,而不是基线唤醒水平。例如,当睡眠或闭着眼静静躺在一个黑暗且安静的房间里时,内向者和外向者的脑活动水平没有不同。当给予中等程度的刺激时,内向者表现出了比外向者更强的生理反应。
设想让内向者和外向者同时执行一项单调的任务,如盯着计算机显示器监视一个核电站的运行情况。显示器很少变化,因此刺激水平很低,工作环境相当枯燥乏味。按照艾森克的理论,在此种情境下,内向者能保持更高的警觉性,工作表现也较好;而外向者的唤醒水平相对较低,很容易昏昏欲睡。然而,假如核电站出现紧急故障,警笛响起,警灯闪烁,人们奔走呼喊……在这种高唤醒条件下,外向者将表现得更好一些,因为内向者会对此刺激表现出过度警觉。
柠檬汁实验:该实验表明内向者对刺激的反应性比外向者更强。有些老师曾经尝试在课堂上做这个实验,结果导致课堂秩序有点混乱。所以,最好的方法是在想象中完成实验,以指出不同类型人格(内向一外向)的不同反应性。实验程序:找一根双头棉签,在正中间系一根细线,调节到悬挂时能使棉签保持平衡(例如,是水平的)。连续吞咽三次后,将棉签的一头放在你的舌头上停留30秒。拿开棉签,滴4滴柠檬汁在舌头上。再做三次连续吞咽,然后把棉签的另一头放在舌头上保持30秒。拿开棉签后,提起中间的细线看它是否保持平衡。如果你是外向者,棉签很可能会保持平衡,因为柠檬汁没有让你分泌较多的唾液,说明你对刺激的反应性低。如果你是内向者,很可能棉签不再保持平衡,因为柠檬汁使你分泌较多的唾液,使放在舌头上的棉签变重了。这就是艾森克所做的经典实验。
该理论的一个重要推论是,当可以自己选择时,外向者选择的刺激水平高于内向者。有间接的证据来支持这个推论。例如,实验室研究表明,当按按钮能够引起视觉环境改变时(如换电视频道,切换幻灯片等),外向者按按钮的频率高于内向者。在大学图书馆里进行的一项自然情境研究中,外向者更多地选择在吵闹的而不是安静的阅览室学习。这些研究表明,当自由选择时,较之于内向者,外向者倾向于选择更高水平的刺激。
心理学家吉恩用一个精巧的设计检验了以下假设:虽然内向者比外向者选择的刺激水平低,但二者在自己选择的刺激水平下,其生理唤醒水平却是相同的。可是,当给外向者呈现内向者选择的刺激水平时,外向者应该会处于低唤醒、疲惫状态,在任务中的表现也会很糟糕。反之,当给内向者呈现外向者选择的刺激时,内向者应该处于过度唤醒的、不适的状态,任务中的表现同样也很糟糕。该假设很复杂,若想了解该研究请参阅后面的阅读材料。
对奖赏与惩罚的敏感性
格雷提出了另一个有影响力的人格生理理论,被称为敏感性强化理论。基于动物脑功能的研究,格雷建构了一个人格模型,其基础是假设人脑内存在两种生理系统。一种是行为激活系统(BAS),它对奖励信号敏感,控制着趋近行为。当BAS意识到某个刺激代表一种潜在的奖励时,就启动趋近行为。例如,在还是孩子时你可能已经知道,当某种音乐响起时就是冰淇淋车在给邻居派发冰淇淋。所以,当听到这个音乐时(奖励线索),你的BAS就产生一种冲动,想要冲到街上找那辆冰淇淋车(趋近动机)。格雷(1975)提出的另一个系统是行为抑制系统(BIS),它对惩罚、失败和不确定的线索敏感,其作用是消除、抑制某种行为或产生回避行为。接着上面的例子,你可能曾经因为冲上街寻找冰淇淋车而受到母亲的批评和惩罚,此时街道变成一个惩罚的线索,使你抑制冲上街道的行为。打个比方,BAS是趋近行为的加速器;而BIS是抑制行为或使人停止行动的制动器。
阅读
吉思的研究
吉恩依据个体对艾森克人格量表中的外向性量表(表2列出了题目)的回答选择被试。30名高分被试组成外向组,30名低分被试组成内向组。要求被试在同一时间段来实验室,并告诉他们将参加“噪音对学习影响”的实验。要求每个被试都完成一项较为困难的联想—配对学习任务,即呈现一对单词,让被试猜测哪个词是实验者依据某一规则选定的,而且被试必须学会这个规则。主要规则有:“所有的词都是动物”,“所有的词以元音字母开头”或“所有的词是颜色的名称”。在被试完成任务的过程中,记录他们的心率和皮肤导电性。
实验开始之前,告知被试在从事作业任务时,会听到来自耳机内的随机噪音。三分之一的被试(内向和外向各10人)可以选择噪音背景的水平。自由选择组的被试可以通过旋钮调节噪音的大小。他们的指导语是调整音量,直到噪音达到“最适合”完成任务的水平。虽然有两个被试(都是内向者)在给出指导语之前要求不要给予噪音,但是还是告知他们不能选择静音作为噪音背景。
该研究中有两个控制组。一组称做相同分配组:三分之一的内向者和外向者分别接受第一组中内向者和外向者选择的噪音水平。另一组被称做交叉分配组:最后三分之一的内、外向者分别交叉接受第一组中外向者和内向者选择的噪音水平。这种实验条件下的被试必须在与自己人格相对的被试选择的噪音背景下完成任务。两个控制组使得实验更有说服力。
关于自由选择组被试选择的噪音强度,结果正如预期的那样,外向者选择的噪音水平显著高于内向者。外向者选择的平均噪音水平是72分贝,内向者的是55分贝。当在自选的噪音水平下工作,或是处在相同分配组的条件下,内向者与外向者之间的反应没有差异。
然而,当采用交叉噪音分配时,两组被试的人格差异就非常明显了。在此条件下,内向者比外向者表现出更高的唤醒水平。当给予内向者选择的噪音水平时,外向者的唤醒水平达到最低,事实上他们感到很无聊;而当给予外向者选择的噪音水平时,外向者的唤醒水平提高,内向者提高得更加明显。外向者觉得“恰好”时,内向者已经是过度唤醒了。
至于作业任务的成绩,内向者被给予外向者选择的噪音时,成绩最差。在外向组选择的噪音条件下,内向者平均需9.1次才能学会联想规则;而在内向组选择的噪音较小的条件下,平均只需5.8次。内向者作业成绩的下降可能是较大的噪音带来过度刺激的缘故。另一方面,外向者在噪音条件下表现得相当好,平均只需5.4次就学会了规则;而当在内向者选择的噪音较小的条件下时,外向者表现得有点糟糕,平均需要7.3次。
这个研究的重要性在于它清楚地证明了,外向者比内向者更偏好高强度的刺激。外向者觉得正好的刺激,对内向者来说已是过度唤醒了,会导致成绩较差。同样,内向者觉得正好的刺激,则会降低外向者的唤醒水平和作业成绩。当分别给予合适强度的刺激时,内向者和外向者才有最好的行为表现。
格雷认为,人们之间的差异在于他们的BAS或BIS的敏感性相对不同。BIS敏感者对惩罚、失败或新奇事物反应强烈,对消极情绪敏感,如焦虑、害怕和悲伤等。BIS负责焦虑这一人格维度。另一方面,BAS敏感者对奖励反应强烈,对积极情绪敏感,倾向于接近刺激。BAS敏感的个体在趋近目标时,抑制行为的能力降低。BAS负责冲动性这一人格维度。
格雷和其他人把冲动性和焦虑模型当做对艾森克的外向性和神经质维度的另一种解释,如图2所示。在格雷的模型中,焦虑和冲动性维度旋转30度即可得到外向和神经质维度。高度外向和轻微神经质的人最具冲动性,冲动性维度的另一端是内向和情绪稳定者。轻微内向和高度神经质的人最容易焦虑,焦虑维度的另一端是外向和情绪稳定者。
在艾森克的外向性和内向性维度所界定的概念范畴中,BAS(冲动性)和BIS(焦虑)确切处在哪个位置还存在一些争议。事实上,本书的作者之一就此问题与格雷及其同事进行了一系列交流。格雷的模型和艾森克的模型之间的关系似乎是直接的:BAS相当于外向性,而BIS相当于神经质。实际上,在前面提到过的坎利等人的研究(2001年)中发现,同内向者比较,外向者对引发高兴和奖励的图像更敏感。而且,与低神经质的人相比,高神经质者的大脑对与负性情绪相关联的图像更敏感。许多研究者认为BIS和BAS与神经质和外向性是类似的,因为它们都涉及到逃避惩罚或追求奖励的一种先天倾向。为此,格雷已经修正了他的模型,对BIS的定位更接近神经质,对BAS的定位更接近外向性。
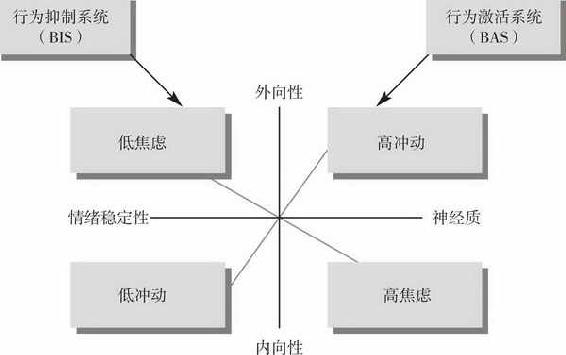
图2 艾森克的外向性和神经质维度与格雷的冲动性和焦虑维度的关系。
格雷相信,对奖励和惩罚的敏感性的不同使人们产生了各种与焦虑/神经质和冲动/外向性有关的行为。如果我们问,为什么有些人比其他人更容易焦虑、害怕、担忧、忧郁、恐惧、困惑或强迫?格雷的回答是,他们具有过于敏感的行为抑制系统。这些人倾向于注意惩罚和失败,且对此非常敏感。此外,他们会因为不确定和陌生而感到压力。那么,如果我们问,为什么有些人比其他人更易受到积极情绪的影响、有更多趋近行为、追求社交?格雷会说这些人具有过于敏感的行为激活系统。
受到格雷理论的启发,一个研究团队构建了测量BIS敏感性的量表。BIS的敏感性是指焦虑、害怕,避免不确定性和避免冒险的倾向。研究者找到了害怕和不害怕两组被试,以确定哪些问题能把他们区分开。其中一些问题是:“我很独立,不受家庭的约束”;“我十分自信”;“有些人很容易被利用,但对于利用者我是不会谴责他们的。”对于高BIS组,研究者选择了一些有焦虑和恐慌发作病史的女精神病患者;而低BIS组需要不顾及自身安危、爱冒险、不考虑危险后果的被试。因此,为了使样本具有代表性,研究者选择了一些有犯罪前科的妓女(经常从事非法活动、高风险的性行为和吸毒)。结果发现,问卷题目能明确区分这两组被试,妓女的分数低于焦虑女病人的分数。该发现说明此问卷在测量对风险环境、危险和无畏的容忍性上具有一定效度。
心理学家卡弗及其同事组成了应用格雷理论的另一个研究团队。他们发展和确定了测量BAS和BIS的量表,随后其他研究者对此进行了验证。例如,有研究者发现这个量表是测量BAS和BIS的最好工具之一。卡弗等人(1999年)对格雷的理论进行了综述,强调个体在趋近或激励动机(外向性或冲动性)上的个体差异,以及在退缩或厌恶动机(内向性或神经质)上的差异。他们将一些研究整合起来,发现人类似乎拥有两个分离的系统分别对诱因和威胁做出反应。这两个系统表现出稳定的个体差异;它们与大部分情感特质相关;都位于大脑皮质的内侧面;对奖励学习和惩罚学习的反应也不同。卡弗和他的同事们将之称为“大二”人格维度。这篇综述显现了格雷人格理论非凡的整合能力。
格雷主要采用动物进行实验,因为对动物可以使用药物或手术来切除某个脑区,然后观察脑区切除后是否会影响到动物的惩罚学习或奖励学习能力。格雷将焦虑和冲动性与两种学习原理联系起来:强化(正强化和负强化)和惩罚(无强化)。许多证据表明这两种学习受不同的神经通路控制。当一个人或一只动物通过奖励或惩罚进行学习时,可能具有不同的脑机制。因此,人们对奖励和惩罚应该具有不同程度的敏感性(高度、中度、低度)。
在一项研究奖励和惩罚的实验中,要求被试完成数百次困难的反应时任务,他们要尽可能快速且准确地说出从电脑屏幕上弹出的词的颜色。这是一项困难的任务,因为被试需要在不到一秒的时间内做出反应,所以只能正确地完成一半任务。对其中一组被试,在每次正确且快速的反应之后给予奖励,他们在20分钟实验时间内能赚取5美元;对另一组被试,在每次错误且缓慢的反应后给予惩罚,尽管在实验开始之前他们曾得到10美元,但实验过后只剩下5美元。这样,两组被试实际上最终都得到了5美元。但其中一组是每次给予奖励,而另一组是每次给予惩罚。结果,BAS的分数能够较好预测被试在奖励条件下的成绩。高BAS者在奖励条件下变得越来越准确和迅速。另一方面,BIS分数能够较好预测惩罚条件下的成绩。与低BIS者相比,高BIS者在惩罚条件下表现得更好。
对格雷理论进行验证的许多研究集中在冲动性(抑制反应的能力)上。我们的监狱里到处是缺乏行为控制能力的人,特别是对于那些能产生即时回报的行为,更是缺乏控制力。例如,一个17岁的男孩看见一辆昂贵的跑车停在路上。他可能会想,要是能开开这辆车该多么爽啊!他注意到车钥匙留在启动装置上,街道上没有人,车主也不在旁边。于是,他开始把手伸向车门……在能得到即时回报的情况下,能否停止趋近行为,是区分冲动者和一般人的依据。
冲动性个体的特征是有着较强烈的趋近倾向而不是退缩行为,他们很少能抵制趋近行为,特别是当趋近的目标或奖励就在眼前时。你可能认识这样的人,他们时常祸从口出或未经思索就伤害了他人的感情。虽然知道自己会伤害他人的感情,也觉得这很糟糕(即受“自责”惩罚)。可是,他们就是不能控制自己的言行,这是为什么呢?
按照格雷的理论,冲动的人惩罚学习的效果较差,因为他们的行为抑制系统较薄弱。如果此理论成立,研究者应该可以证明,在从事包含惩罚的任务时,冲动者比非冲动者的表现要差。分别对冲动性的大学生、少年犯、精神病患者和在押犯人进行的研究发现,这些人确实缺乏惩罚学习的能力。例如,当冲动者玩游戏时,与因正确反应而获得奖励相比,因错误反应而受罚时,他们学得更慢。似乎冲动性的人惩罚学习的效果不如奖励学习的效果好。
假设你有一个室友,你想让她养成打扫室内卫生的习惯。当她每一次收拾屋子时,你可以尝试用糖果或表扬来给予奖励;或者当她每一次乱丢乱放时,你用尖叫或指责来给予惩罚。如果她是个冲动的人,你最好采用奖励策略;相反,如果她是个焦虑的人,你最好采用惩罚策略。
假设有这样一个情境:你打算教他人学习一些新东西。先举例讨论你会采取何种奖励策略教他(她),再接着讨论你会采取何种惩罚策略教他(她)学习同样的内容。
感觉寻求
另一个被认为存在生理基础的人格维度是感觉寻求。感觉寻求是指一种寻求刺激和兴奋、享受冒险、避免厌倦的倾向。对此维度的研究源于对感觉剥夺的研究。所以,我们将从感觉剥夺的研究开始阐述。
假设你自愿参加一项研究,研究中你被关进一个很小的房间。房间里没有灯光,没有声音,仅保留最微弱的触觉。假设你同意在里面待够12个小时,那么这将是一种怎样的感觉?研究表明,一开始你会感到放松,接着觉得无聊,之后开始焦虑,以致出现幻觉和错觉。赫布(1955)的早期研究显示,在这种情境下,大学生宁肯选择反复听一盘告诫6岁男孩不要喝酒的磁带。在早期的感觉剥夺实验中,还有些志愿者选择一遍遍地听一盘老式的股市报告录音,以避免感觉剥夺的不愉快体验。在感觉剥夺实验中,被试似乎愿意寻求任何形式的感觉输入,即使是那些在正常情况下非常无聊的刺激。

感觉寻求理论解释了为什么尽管有些刺激体验会带来特定的风险,但是一些人仍然乐此不疲。
赫布的最佳唤醒水平理论
赫布提出了最佳唤醒水平理论,它被艾森克用于其外向性理论之中。赫布认为,个体试图达到最佳唤醒水平。如果低于最佳唤醒水平,唤醒水平的增加就是一种奖赏;反之,如果高于最佳唤醒水平,那么唤醒水平的降低就是一种奖赏。在赫布那个年代,其理论是充满争议的。因为大多数研究者认为缓解紧张是所有动机的目标。而赫布却指出,我们的动机是寻求紧张和刺激。我们该如何解释这样的事实:人们喜欢玩智力拼图;喜欢轻微的挫败感;偶尔冒险或做一些引起轻度害怕的事情,如坐过山车。赫布相信个体需要刺激和感觉输入,这与感觉剥夺的研究一致。而且,至少神经系统本身需要一些感觉输入。
查克曼的研究
查克曼和哈伯注意到,在早期的感觉剥夺研究中,有些人经历感觉剥夺后并不像其他人那么痛苦;而有些人却觉得感觉剥夺特别痛苦,整个实验期间他们不断要求放录音或阅读材料,退出实验的时间也相对较早。查克曼认为,这些人对感觉剥夺的耐受性低,对感觉刺激的需求高。他称这样的人为感觉寻求者,因为他们不仅在感觉剥夺实验里,而且在日常生活中也总是倾向于寻求刺激。
查克曼编制了一个量表,用来测量人们需要新奇或兴奋体验和醉心于激动或兴奋的程度。他称此量表为感觉寻求量表,表3列出了该量表的部分项目。查克曼假设,一些人(高感觉寻求者)需要大量刺激以达到最佳唤醒水平。此外,当刺激和感觉输入被剥夺时(如关在一个感觉剥夺的小房间里),这些人会觉得特别难受。
果不其然,查克曼编制的测量人们日常生活中刺激偏好的量表可以很好地预测其忍受感觉剥夺的程度。高感觉寻求者认为感觉剥夺实验特别难受;而低感觉寻求者则能忍受更长时间的感觉剥夺。20世纪60年代早期,查克曼放弃了对感觉剥夺的实验室研究,转而开始了对与感觉寻求维度有关的其他特征的研究。注意,查克曼对感觉寻求的理论诠释与艾森克对外向性的解释是一致的。事实上,感觉寻求和外向性之间具有中等程度的正相关。
表3 感觉寻求量表的项目

感觉寻求量表的所有题目及记分方式,可参看查克曼(1978)的量表。
30多年来,查克曼和他的同事,以及其他研究者致力于感觉寻求的研究,得出了许多有趣的发现。大量研究一致发现:高感觉寻求者在日常生活中需要高水平的刺激。在感觉寻求量表的得分上,自愿参加平暴行动的警察比不愿参加平暴行动的警察要高;跳伞者的得分比不跳伞者高。在自愿参加心理实验的大学生中,感觉寻求得分较高者更喜欢非常规的研究(如ESP、催眠或吸毒的研究),而不是经典研究(如学习、睡眠或社交研究)。在赌博行为的研究中,高感觉寻求者更喜欢下大注。根据报告,高感觉寻求者拥有大量性伙伴,喜欢各种性行为,比低感觉寻求者更早发生性行为。
查克曼认为感觉寻求具有生理基础。他最近的研究(1991年)主要聚焦于神经递质在感觉寻求差异中的作用。神经递质是神经元内的化学物质,其作用是在神经元之间传递神经冲动。你可能记得曾在心理导论课上学到,相邻神经元之间有一个很小的间隙(称为突触间隙),神经冲动要持续传递到达目的地,就必须穿过这个间隙。神经递质是神经元释放的化学成分,它使得神经冲动通过突触继续传递下去。
一旦神经冲动传导完成,神经递质将被分解。否则,就会导致过多的神经传导。就像剧院或地铁的旋转门,一次只允许一个人进入。如果把门打开,许多人都可以通过,这会使太多人进入;如果门紧闭着,那么就没人能够进入。神经递质系统与此类似,突触内的化学平衡必须适当,从而使适量的神经传递正常通过突触,继续进行传导。
某些酶,特别是单胺氧化酶(MAO)能够使神经递质维持在合适的水平。当一种神经冲动通过后,单胺氧化酶就会将多余的神经递质分解。如果单胺氧化酶过多,就会导致太多的神经递质被分解,神经传导将受阻;如果单胺氧化酶过少,会使太多的神经递质堆积在突触内,以致发生过多的神经传导。假设你要用手指做一个简单的动作,如从平坦的地面捡起一枚硬币。如果你体内的单胺氧化酶太少,你的手指会颤抖,动作急促(太多神经传导);然而,如果单胺氧化酶太多,你的手指将会很笨拙,因为你感觉迟钝、运动控制感匮乏。只有当单胺氧化酶适量,神经递质维持在恰当的水平时,神经系统才能正常地控制肌肉、思想和情绪。
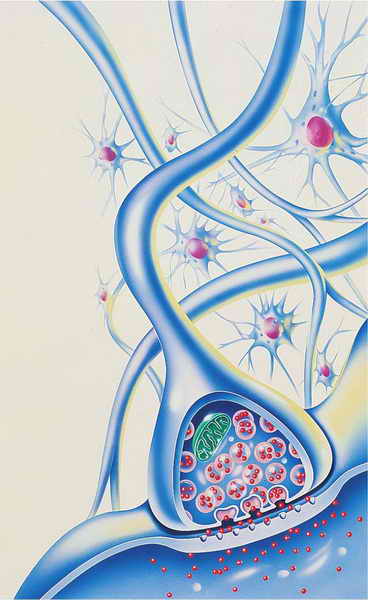
两个神经元的连接处就是一个突触。突触将电信号从一个神经元传递给下一个神经元。当一个电信号到达突触时,就引起了突触小泡(粉色)内神经递质(红色)的释放。突触小泡的隔膜破裂,形成一个微小的间隙,称为突触间隙。神经递质通过这个间隙与受体结合,从而引起了电冲动的传导。
与低感觉寻求者相比,高感觉寻求者血液里的单胺氧化酶水平较低。交叉研究表明,感觉寻求与单胺氧化酶水平之间存在着从低至中等水平的相关,且均为负相关。如果高感觉寻求者的单胺氧化酶水平倾向于较低,而单胺氧化酶低意味着神经元内有更多的神经递质,那么感觉寻求的产生和维持或许是因为神经系统内神经递质过多的缘故。单胺氧化酶就像神经系统的制动器,通过降解神经递质来抑制神经传导。单胺氧化酶水平低,感觉寻求者神经系统的抑制水平低,因而对行为、思想和情绪的控制程度低。依据查克曼(1991)的理论和研究,感觉寻求行为(如不恰当的性行为、吸毒和疯狂派对)并非是由寻求最佳唤醒水平引起的,而是因为神经突触内的生化“制动器”(MAO)太少。
神经递质与人格
查克曼的理论关注单胺氧化酶(可以分解神经递质)的水平,而其他研究者假设,神经递质本身的水平决定了特定的个体差异。神经递质作为人格差异的一种可能来源,已经受到了广泛关注。神经递质中的一种——多巴胺,似乎与高兴相关。例如,为了得到多巴胺,动物会像渴望得到食物那样工作。因此,多巴胺似乎起着奖赏的功能,甚至被称为快乐剂。成瘾物质,如可卡因,作用机理类似神经系统中的多巴胺。这就是摄入可卡因会使人快乐的原因。但是,这些药物降低了人体内自然分泌的多巴胺水平,当神经系统内的药物消失后,就会使人产生厌烦的情绪,从而驱动人急切地获取更多的药物。
第二种重要的神经递质是5-羟色胺。有研究者曾撰文指出5-羟色胺在抑郁及其他心境障碍(如焦虑)中的作用。有些药物,如百忧解、佐洛夫和帕罗西汀妨碍了5-羟色胺的再吸收,使它们长时间滞留在突触内,从而缓解抑郁者的抑郁症状。在一项研究中,让非抑郁的被试服用百忧解。经过几周观察发现,与控制组被试相比,这组被试报告的负性情绪更少,外出和社交行为更多。在以猴子为被试的研究中,5-羟色胺水平高的猴子控制欲较高,更喜欢修饰自己;而5-羟色胺水平低的猴子则更频繁地表现出害怕和攻击行为。综合动物研究的结果,有研究者提出,低5-羟色胺水平与易怒行为相关。
第三种重要的神经递质是去甲肾上腺素,它是交感神经系统对抗—逃避机制的激活剂。无疑,许多人格理论是基于多巴胺、5-羟色胺和去甲肾上腺素等神经递质提出的。也许最全面的是克罗宁格的三维人格模型。该模型认为,有三种人格特质取决于这三种神经递质的水平。第一种特质是新异寻求,以低水平多巴胺为基础。回想一下,低水平的多巴胺使个体迫切想要获取能够增加多巴胺的物质或经验,而新异、刺激、兴奋可以补偿过低水平的多巴胺。因此,新异寻求行为被认为是由多巴胺水平过低引起的。
克罗宁格模型中提到的第二种人格特质是伤害避免,与5羟色胺的异常代谢有关。虽然其他理论认为5-羟色胺的增加或减少与伤害避免有关,但克罗宁格本人(与作者的个人交流,2003年10月)认为,5-羟色胺的绝对水平和伤害避免之间存在简单线性相关的说法是不明智的。脑脊液中的5-羟色胺的主要代谢产物——5-HIAA的水平很低时,存在严重抑郁的风险。但在焦虑和压力状态下,5-羟色胺的水平却能升高。选择性5羟色胺对抑制剂的吸收(如抗抑郁药百忧解、佐洛夫和帕罗西汀)能引起突触间隙内的5-羟色胺水平升高,从而使人首先产生焦虑感,然后减少对压力的过度反应。这可能是因为应对压力时释放的5-羟色胺降低了其敏感性。因此,我们必须明确5羟色胺的重要作用:在急性应激状态下,它会升高;在整个生命过程中5-羟色胺水平的降低与低水平的避免伤害有关。伤害避免水平较低的人表现为充满活力、开朗、乐观;而伤害避免水平较高者则表现为警觉、压抑、羞怯和不安。伤害避免水平高的人似乎总是预期伤害性的、不快乐的事情即将发生,所以他们不停地寻找威胁性的信号。就像狗因害怕狂吠,而不是愤怒一样,伤害避免水平高的人是易怒、敏捷和有敌意的。
第三种人格特质是奖励依赖,克罗宁格认为它与低水平的去甲肾上腺素有关。奖励依赖水平高的人坚持不懈,持续地以能带来回报的方式行事。他们能千方百计、连续数小时不停息地工作。即使其他人都放弃了,他们还在继续奋斗。
基因通过神经递质系统影响人格
虽然我们将在随后的章节里更详细地讨论行为遗传学的内容,但这里值得一提的是,许多对基因和人格感兴趣的研究者把研究重点放在了基因对神经递质的调节上。例如,如果低水平的多巴胺与新异寻求相关,那么,负责多巴胺传导的基因可能是研究人格特质遗传基础的好的着眼点。芬兰的研究者们发现,多巴胺D4受体基因(D4DR)与高水平的新异寻求有关。但是,其他的研究没有发现相同的结果。一项对新异寻求的元分析研究发现,D4DR基因上的一些特定遗传密码确实与新异寻求有关。这些发现意味着,任何一种人格特质的形成都可能涉及多种基因。因此,寻找人格特质的某种基因基础就像大海捞针一样。现在的研究者开始寻找人格特质的多种基因基础,也就是说,他们在寻找多种基因以交互作用的方式对神经递质系统产生的影响。该领域的权威研究者迪安哈默最近发表评论指出:“非常明显,10年后,至少可以发现大部分人格特质与大量基因有关”。随着基因序列分析这一新技术的发展,此类研究将变得更易于操作。然而,将来的研究可能会发现:对于任何具有生理基础的人格特质来说,基因之间复杂多样的交互作用可能需要环境来引发。
克罗宁格的理论对精神病学也有一定的影响,它可用来解释不同类型的成瘾。例如,酗酒者并非都因相同的原因而成为酒鬼。克罗宁格认为,有些酗酒者开始饮酒是因为其新异寻求感较高。他们饮酒是为了补偿过低的多巴胺水平,享受多巴胺升高带来的快感。另一些酗酒者开始饮酒是因为他们是高伤害避免者,是为了缓解长期的压力和焦虑感。他们饮酒的主要动机是解除焦虑。理解人们物质滥用的动机可以帮助他们克服成瘾。例如,一些人吸烟是因为吸烟能缓解压力,而另一些人吸烟的原因在于吸烟能增强活动的愉悦性,就像喝咖啡与社交活动。
很明显,克罗宁格的理论与格雷、艾森克和查克曼的理论有许多共同点。例如,新异寻求与格雷理论中行为激活系统的奖励敏感性相似。但是,这些理论对人格的生理基础有不同的解释。例如,格雷认为,决定人格特质的重要的生理基础是控制奖励学习和惩罚学习的大脑系统。艾森克也强调脑和神经系统的作用。查克曼聚焦于神经突触和在神经突触中发现的神经化学物质。克罗宁格则特别强调特定的神经递质。所有这些理论描述的或许是同一行为特质,只是站在不同的生理角度(从突触到大脑)进行解释。
现在让我们来看看另外两种人格维度,它们的生理基础似乎与生理反应无关。这两种人格维度是早晨型—夜晚型和大脑的不对称性。
早晨型—夜晚型
你也许是个喜欢晚睡晚起的人,将重要的学习或工作任务放在午后或晚上来做,因为那时你精力最旺盛。或许你是个早晨型的人,生活很有规律,不用闹钟每天也能很早起床。你倾向于一大早做重要的工作,因为这个时候你感觉状态最好,并且晚上睡得也很早。早晨型和夜晚型的人表现出稳定的特征。人格心理学家对这种稳定的差异很感兴趣,用早晨型—夜晚型来描述这一人格维度。
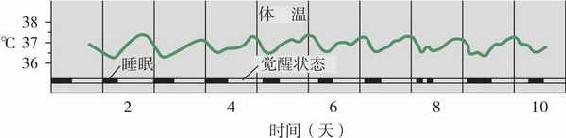
图3 体温的昼夜节律
早晨型—夜晚型的人格差异有时也被称为“百灵鸟型”和“猫头鹰型”。二者差异的根源在于内在生物节律的不同。许多生理过程呈现出大约24~25小时的波动周期,这就是昼夜节律。研究者特别感兴趣的是体温和内分泌的昼夜节律。例如,一般而言,人的体温在晚上8~9点左右最高,而在早晨6点左右最低。图3显示了一天之中人体体温随时间变化的情况。
研究者采用时间隔离设计来研究昼夜节律。时间隔离设计是这样的:被试自愿住进一个由实验者完全控制时间线索的环境里。房间内没有窗户,所以被试无法知道是白天还是黑夜。没有固定的进餐时间,被试不知道吃的是早饭、午饭还是晚饭,只要想吃东西就能得到食物。没有现场直播的电视或电台节目,但是被试有大量的影碟或录音带以供消遣。实验时间为几个星期或更长。多数时候,志愿者是一些学生,因为他们需要利用单独的时间准备一项重要的考试或是撰写论文。
假如你是该实验的一个被试,你可以想睡就睡;想睡多久就睡多久;什么时候想吃就吃;只要愿意,随时可以看电影或工作。这被称为自由运转时间,在此期间,没有任何时间线索影响你的行为或生理。在这样的环境下,你的体温每小时被测一次,如果你和大多数人一样,你就会发现你的体温遵循24~25小时的一个波动周期:醒来之前开始升高,睡觉之前开始下降。
值得注意的是,24~25小时的波动周期是平均值,人与人之间生理节律的实际值有较大的差异。采用时间隔离研究得到的昼夜节律中,最短者只有16小时,最长的可达50小时。在时间隔离实验的“自由运转”时间里,最短的睡—醒周期为16小时,而最长的睡—醒周期持续约50多个小时。
如此大的差异只出现在时间隔离研究的实验情境下。现实生活中,我们周围有许多在24小时左右波动的时间线索,其中最显著的当数昼夜交替。这些时间线索使我们适应了一天24小时的周期。尽管生理节律高于或低于24小时的人对24小时周期适应得较好,但他们生理节律的峰值与谷值存在差异。假设一个人的昼夜节律稍长,约26小时,而另一个人的较短,为22小时。两个人都需要适应一天24小时这一周期,但是前者体温的峰值可能出现得相对较晚(约晚上10点左右),后者的体温峰值出现得相对早些(约为下午6点)。
生理节律短的个体体温和警觉水平的峰值在一天之中出现的早一些,因此,他们比那些生理节律长的人睡得早。虽然闹钟已经告知新的一天开始了,但生理节律为26小时的人很难在早晨6点起床,因为还差2小时才到他们的周期。生理节律为22小时的人很早就能起床,因为他们已经完成了一个22小时的生理节律,在24小时的时钟信号发出之前就已经开始新的一天了。
你认识早晨型的人吗?有什么具体的依据使你断定他(她)是早晨型的人?早晨型的人与夜晚型的人还有哪些不同?例如,早晨型的人还有哪些人格特质?本杰明·富兰克林说:“早睡早起可以使人健康、富有和聪明。”你认为早晨型的人确实更聪明,或是在生活中能够取得更好的成就吗?怎样设计一个研究来回答这个问题?
对昼夜节律的个体差异的研究为理解早晨型—夜晚型的人格奠定了基础。你已经知道,生理节律短的人多是早晨型,而生理节律长的人多是夜晚型。霍恩和奥斯伯格编制了一个包含了19个题目的量表来测量早晨型—夜晚型人格(表4)。题目主要涉及个体在一天里喜欢什么时间活动。在一项研究中,研究者招募了48名被试,连续数天里每隔一个小时对其体温进行测量。结果发现,他们在此量表上的得分与每天体温达到最高的时间之间的相关系数为-0.51。该研究最早是在瑞典进行的,随后在美国、意大利、西班牙、克罗地亚和日本进行的研究也得到了同样的结果。
表4 早晨型—夜晚型人格问卷的题目
表4 续表
表4 续表
这些得到跨文化研究重复验证的结果符合早晨型—夜晚型活动的理论。一天中的最佳时间是一种稳定的、具有生理基础的特质。运用霍恩和奥斯伯格编制的量表测量的结果也具有跨时间的稳定性。克罗地亚的研究者用此量表测试了90名大学生,待7年后他们大学毕业时进行重测。结果发现两次测量结果呈显著的正相关,说明早晨型—夜晚型人格特征具有跨时间的稳定性。然而,整个样本也表现出向早晨型转变的趋势。可能的原因在于,这是一个从大学生向工作者转移的群体。
许多研究者研究了该量表的效度。例如,在一项研究中,研究者要求大学生被试完成连续84天的自我记录,记录内容包括:每天何时睡觉,何时起床,何时感觉状态最好。霍恩和奥斯伯格的量表与被试的平均作息时间,以及每天感觉最好的时间具有高度正相关。通常,早晨型的人比夜晚型的人起得早、睡得早,最佳状态也出现得早。
如果早晨型和夜晚型的人不得不住在一起(如大学室友)结果会怎么样呢?有人喜欢晚睡晚起,有人则喜欢早睡早起,即使在周末亦如此。你认为他们住在一起会快乐吗?有研究者以密歇根州立大学一年级的住校学生为被试对该问题进行了研究,被试都只有一个室友。室友们除了完成早晨型—夜晚型问卷(MEQ)之外,还对彼此的友谊进行全面评价。结果发现,室友之间MEQ得分的差距越大,友谊评价等级也越低。得分差距较大的室友们说,他们彼此相处得很不好,不喜欢这种关系,彼此不是朋友,也不想继续住在一起。其他人格维度上的差异,如成就动机和竞争性,不能预测室友关系的满意程度。这似乎表明,早晨型—夜晚型的人格差异与人际关系融洽问题直接相关。
其他研究关注这两种类型的人格在一天的不同时段内的认知表现。研究者们先用早晨型—夜晚型问卷对被试进行分类,然后测试他们在一天不同时间里完成系列搜索和逻辑推理任务时的表现。结果发现,早晨型的人在上午8~11点表现得最好,而夜晚型的人在下午5点至晚上11点表现最佳。另一些研究者发现,这种差异可以通过使用兴奋剂(如咖啡因)来减弱。如果在早晨摄入咖啡因,可以大大提高夜晚型的人的表现;如果在晚上摄入咖啡因,则可以大大提高早晨型的人的表现。人们可以根据自己是早或晚的类型,定时摄入咖啡来充分提高工作和学习效率。
早晨型或夜晚型的人偏好一天内的某些时段是有其生理基础的。然而,有时候却会打乱这种偏好。比如一个夜晚型的大学生,他需要选修的课程只被安排在上午8点;或者在工厂上班的一个早晨型工人,被安排上“晚班”(下午4点到午夜)。改变一个人的自然生理节律是困难的,但并非不可能。人们可以适应倒班的工作或者睡—醒周期的改变。有些证据表明,夜晚型的人调节被打乱的睡—醒周期的能力好于早晨型的人。对于穿越子午线飞行(产生飞行时差)或整晚上班不休息(通宵工作)的情况,夜晚型的人的耐受性好于早晨型的人。
总之,人们在一天之中何时开始活跃,喜欢在何时做重要的事,需要早些时候还是晚些时候开工,取决于内在昼夜节律的长短。这是人格生理取向研究的一个典型例子,因为它强调了这样的观点:行为模式(即对一天中不同时段的偏好)取决于内在的生理机制(昼夜节律)。
大脑不对称性与情感类型
你可能意识到左右脑是专门化的,在控制不同心理机能方面具有不对称性。受到研究者关注的一种不对称性是左右脑半球额叶的相对活动量。大脑会持续地产生少量的电活动,我们可以通过装在头部的敏感装置对其进行测量,并称这种电活动的记录为脑电图(EEG)。随着脑的神经生物活动呈现出或快或慢的波动,这种电活动是有节律的。有种脑电波被称为α波,频率为每秒8~12次。某时段α波的数量是该时段脑活动的反向指标。α波产生于个体安静、放松、感到有点困和对环境不加注意的状态下。在一个特定时段的脑电波记录中,α波出现得越少,此时的脑活性越高。
在脑的任何区域都可以测得脑电波。在情绪研究中,研究者特别关注脑的额叶,比较左右脑的活动性。研究结果表明,当人处在高兴的状态下时,左半球比右半球的活动性更高;反之,当人不高兴时,右半球比左半球活动性更高。例如,戴维森及其同事让被试看一些能引起人愉快或厌恶情绪的电影片段,同时记录他们的脑电波,并对他们看电影片段的过程进行录像。结果发现,愉快时,被试的左额叶较右额叶显示了更多的脑活动;反之,当被试表现出厌恶表情时(抿嘴、咂舌、皱鼻),被试的右额叶比左额叶显示了更多的脑活动。
对婴儿的研究也得到了相似的结果。福克斯和戴维森没有使用电影片段,而是将或甜或苦的汁液滴进十个月大的婴儿嘴里,引发其愉快或不愉快的情绪反应。结果,甜汁液诱发了更多的左脑活动,苦汁液则诱发了更多的右脑活动。在对10个月大的婴儿进行的另一项研究中,研究者让婴儿的母亲离开,把婴儿单独留下,接着一个陌生人进入房间。在这个标准的引发焦虑的情境中,一些婴儿变得痛苦,一些婴儿开始哭闹和喊叫,另一些则不然。研究者据此将婴儿分为两组,哭喊组和不哭喊组。结果发现,同不哭喊组相比,哭喊组表现出了更多的右脑活动(相对于左脑)。研究结果揭示,痛苦与否的倾向(与EEG不对称性有关)是婴儿的一种稳定特征。福克斯及其同事对婴儿的额叶脑电不对称性进行了重复测量(分别在7个月和12个月时),两次测量结果之间高度相关,这说明两侧额叶脑活动的不对称性具有跨时间的稳定性。在成人中也得出了类似的结果,各种研究中的EEG不对称性的重测相关在 .66~.73之间。这些发现说明,前额不对称性的个体差异是相当稳定和一致的,可以将它作为内在人格特质的生理指标。
其他研究也揭示出EEG的不对称性是愉快易感性的指标。许多研究者都曾考察过正常被试对情感性电影片段的反应与其前额不对称性之间的关系。在这些研究中,先在被试休息状态下测出EEG的不对称性类型,接着给被试播放好笑、快乐的电影和痛苦、可怕的电影,要求被试回答电影带给他们的感受,作为因变量。研究假设是,在休息状态下右脑活动性更高(看电影之前的测量)的被试对痛苦、可怕的电影会报告更多的负性情绪。与之相对,左脑活动性更高的被试对好笑、快乐的电影会报告更多的正性情绪。预测得到了实验结果的支持,看电影之前的额叶不对称性测量,预示了接下来被试自我报告的对电影的情绪反应。右脑优势的被试对令人厌恶的电影报告了更多的痛苦;而左脑优势的被试对令人快乐的电影有更快乐的反应。
没有EEG也可以评估脑的不对称性。EEG不是获取脑活动不对称性的惟一途径。当人集中精力回答难题时,眼球偏向的方向也可以反映他的左右脑活动水平。例如,要求一个人用“狂喜”和“高兴”造一个句子。在思索时,人的眼球会偏到左边或是右边。对右利手的人来说,偏向右边意味着左脑活性高,偏向左边意味着右脑活性高。如果问一个人几个难题,如“从你的住所走到最近的商店,需要转多少个弯?”你就会注意到他的眼睛通常向一边偏。据此可以判断他的优势脑。当然,这种快速的判断并不如EEG可靠,但还是可以作为一种粗略的测量手段。
你可以找几个朋友或熟人试试这个方法。大多数人眼球偏向的方向不是每次都一致,所以多问几个问题很重要,这样可以观察最常见的方向。观察的同时,你还需要确定他们的情绪通常是正性还是负性。频繁偏向右边的人是左脑优势,应该易于表现出更多的正性情绪(如幸福、愉快、热情);频繁偏向左边的人是右脑优势,应该易于表现出更多的负性情绪(如痛苦、焦虑、悲伤)。
当然,影响人们感受和情绪的因素很多。脑活动方式只是其中之一,它影响了我们体验某种情绪的可能性。
在猴子身上也发现了相似的结果。因为猴子无法告知我们它的感觉是积极的还是消极的,所以研究者通过检测皮质醇来评价猴子的情绪反应。皮质醇是一种应激激素,为机体的对抗或逃避反应做准备。皮质醇含量的增加表明动物最近体验到了压力。戴维森及其同事发现,猴子的右脑越活跃,皮质醇水平越高。最近,在6个月大的婴儿身上也发现了相同的结果。研究者安排一个陌生男人进入房间,慢慢靠近婴儿,盯着婴儿看两分钟,从而引发婴儿的惊恐。此时那些右脑基线水平更活跃的婴儿的皮质醇水平增加。同时,与左脑活跃的婴儿相比,右脑活跃的婴儿表现出更多的哭喊行为、害怕表情和试图逃跑的反应。
戴维森和萨顿发现,具有积极情绪倾向的人(用怀特和卡弗于1994年编制的BIS/BAS量表测量所得)在没有情绪刺激的情况下,左前额的EEG的基线水平相对更活跃。显然他们吸收了格雷的理论,把情感类型和脑功能结合起来,表明了格雷提出的BIS和BAS概念的效用,以及二者不同的激活机制(趋近动机和逃避动机)。最近,有另一批研究者应用功能性脑成像技术验证了这个结果。
大脑不对称研究的重要性在于,给予不同的情绪刺激,大脑的不同区域会对快乐或厌恶情绪产生不同的反应。福克斯和卡尔金斯用反应阈限的观点来解释这个现象。当不快乐事件发生时,右脑活跃的人对负性情绪的反应阈限降低,只需很少的情绪刺激就能唤起他们的负性感受。而左脑活跃的人对正性情绪的反应阈限较低。反应阈限的概念意味着,只需很少的情感刺激,就能诱发出左脑或右脑活跃者相应的情绪反应。脑功能的不对称可能是情感类型的根源,或者至少可以预测人的情感类型。


