
![]()

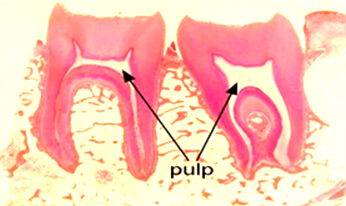
牙髓(pulp)是来源于外胚间叶的疏松结缔组织,位于由牙本质所形成的髓腔(髓室和根管)内。牙髓的主要功能是形成牙本质、营养、感觉、防御及修复。牙髓中的血管、淋巴管和神经仅通过根尖孔与根尖部的牙周组织相通连。

牙髓是疏松结缔组织,它包含有细胞、纤维、神经、血管、淋巴管和其他细胞外基质。牙髓组织可分为四层,即:
◇ 靠近牙本质的一层为成牙本质细胞层;
◇ 紧接着成牙本质细胞层,细胞相对较少的组织为乏细胞层,或称 Weil层 (the zone of Weil),此层约为40μm宽,有血管、无髓神经纤维和纤细的成纤维细胞的突起分布。此层存在与否决定于牙髓的功能状态,在牙本质形成较快和有修复性牙本质形成的区域可能无此层;
◇ 无细胞层内侧细胞密集,称多细胞层;
◇ 牙髓中央区细胞分布比较均匀,称固有牙髓(pulp proper)或髓核(pulp core),含丰富的血管和神经。
【细胞】
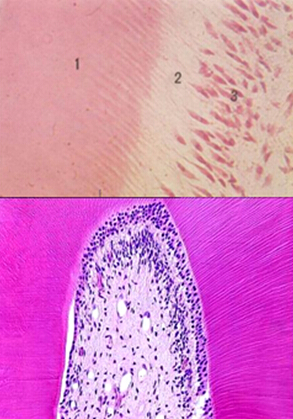 ◇ 成牙本质细胞(odontoblast),位于牙髓周围,呈柱状紧接前期牙本质排列成一层,是呈极性分布的终末分化细胞。其细胞顶端有一细长的突起伸入牙本质小管内,因此成牙本质细胞层实际上由成牙本质细胞的胞体构成。成牙本质细胞排列成栅栏状,并且细胞彼此拥挤,细胞核并不在同一水平,在光学显微镜下,似由3~5层成牙本质细胞构成。在整个牙髓中,成牙本质细胞的形状并不完全一致,在年轻恒牙的冠部为较高的柱状细胞,反应了细胞的高活性状态;在牙根中部逐渐变为立方形细胞;接近根尖部的造牙本质细胞为扁平状,呈现相对休止状态。在成牙本质细胞之间,有时可见毛细血管、神经纤维和树枝状细胞分布。
◇ 成牙本质细胞(odontoblast),位于牙髓周围,呈柱状紧接前期牙本质排列成一层,是呈极性分布的终末分化细胞。其细胞顶端有一细长的突起伸入牙本质小管内,因此成牙本质细胞层实际上由成牙本质细胞的胞体构成。成牙本质细胞排列成栅栏状,并且细胞彼此拥挤,细胞核并不在同一水平,在光学显微镜下,似由3~5层成牙本质细胞构成。在整个牙髓中,成牙本质细胞的形状并不完全一致,在年轻恒牙的冠部为较高的柱状细胞,反应了细胞的高活性状态;在牙根中部逐渐变为立方形细胞;接近根尖部的造牙本质细胞为扁平状,呈现相对休止状态。在成牙本质细胞之间,有时可见毛细血管、神经纤维和树枝状细胞分布。
成牙本质细胞的主要功能是形成牙本质(包括牙本质中的纤维、基质和牙本质的生物矿化)。在正常情况下只要牙髓保持活力,牙本质在牙的一生中都可形成。
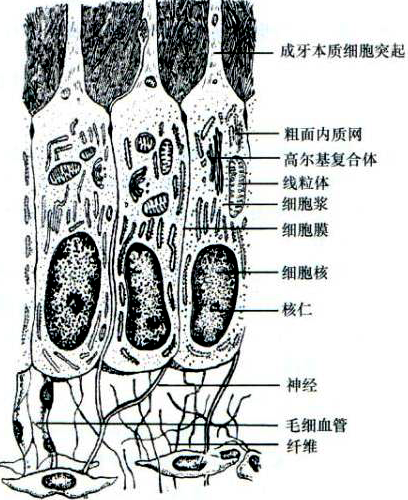
电镜下,成牙本质细胞的胞核位于远离其突起的基底部,核的上方有粗面内质网和高尔基复合体。胞质内线粒体和溶酶体散在分布,还可见其他成分如细胞骨架和分泌泡。在牙本质形成活跃期,细胞内高尔基复合体显著,粗面内质网丰富,线粒体遍布于整个胞质内。
相邻的成牙本质细胞间有特化的细胞间的连接即连接复合体,包括桥粒、缝隙连接和紧密连接。位于胞体顶端(近突起处)呈斑状分布的桥粒形成成牙本质细胞间的机械结合。大量的缝隙连接提供了细胞间信号分子通道,以维持统一的细胞活动,形成均匀一致的前期牙本质层。原发性牙本质形成时,这种连接复合体最多。成牙本质细胞与其下层的成纤维细胞间也有桥粒和缝隙连接。紧密连接主要见于年轻牙的成牙本质细胞顶端。这些结构由细胞间线性的嵴或沟形成,关闭细胞间隙。牙本质被釉质和牙骨质覆盖后,紧密连接似乎可确定成牙本质细胞层的渗透性,可限制各种分子、离子和液体在牙髓和前期牙本质间通过。窝洞制备时这些连接被破坏,牙本质的渗透性增加。
成牙本质细胞合成和分泌的蛋白质主要是Ⅰ型胶原,也有少数Ⅴ型胶原,非胶原成分包括蛋白多糖、磷蛋白、糖蛋白、含γ羧基谷氨酸蛋白等。成牙本质细胞中也存在其他可溶性分子如TEFβ、EGF、FGF,这些分子可能在创伤愈合中起重要作用。在成牙本质细胞内还有对牙本质矿化起重要作用的牙本质磷蛋白(DPP)和牙本质涎蛋白、牙本质基质蛋白、核心蛋白聚糖(decorin)和双糖链蛋白聚糖(biglycan)。这些分子都具有特殊的功能,如双糖链蛋白聚糖和Ⅰ型胶原分泌在成牙本质细胞――前期牙本质界面,在此处双糖链蛋白聚糖连接胶原,在组成胶原纤维中起作用;核心蛋白聚糖和DPP分泌在矿化前沿即矿化牙本质和前期牙本质界面,在此处核心蛋白聚糖组织和搭配胶原纤维,以利于DPP与钙结合而启动牙本质矿化。其他在牙本质细胞外基质中的物质也由成牙本质细胞合成,如各种糖胺聚糖、胶原酶及其组织抑制物、糖蛋白如骨钙素、骨桥蛋白等。
-----------------------------------------------------------------------------------------
◇ 成纤维细胞,是牙髓中的主要细胞,故又称为牙髓细胞。细胞呈星形,有胞质突起互相连接,核染色深,胞质淡染、均匀。电镜下有丰富的粗面内质网和线粒体以及发达的高尔基复合体等,这说明它有活跃的合成胶原的功能。主要合成Ⅲ型胶原和其他细胞外基质如蛋白多糖和糖胺聚糖,成纤维细胞也能降解细胞外基质。牙髓成纤维细胞的形态往往反应了牙髓组织的功能和活性。随着年龄的老化,牙髓成纤维细胞数量减少,形态呈扁平梭形,细胞器减少,表现为合成和分泌功能下降。
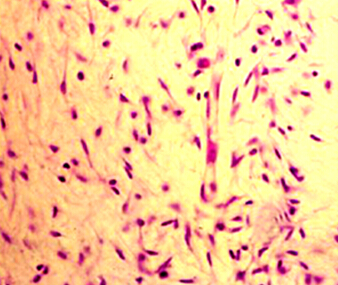
成纤维细胞在创伤修复机制中的作用非常重要。在适当的刺激下如暴露的前期牙本质或炎症细胞释放的生长因子、某些骨形成蛋白、细胞因子或炎症介质的刺激,成纤维细胞可增生、分化为新的成纤维细胞或成牙本质细胞。
-----------------------------------------------------------------------------------------
◇ 巨噬细胞和未分化间充质细胞(macrophages and undifferentiated mesenchymal cell) 这些细胞通常位于小血管及毛细血管周围。巨噬细胞为较大的椭圆形或梭形,胞核染色深。在活体染色法中,可见其胞质内储有染料颗粒。电镜下胞质含溶酶体。在非活动时期很难与成纤维细胞相鉴别。巨噬细胞在牙髓成纤维细胞更新能吞噬死亡细胞,也在炎症时发挥作用。
以前,能形成第三期牙本质的细胞被冠以许多名称如未分化间充质细胞、多潜能间充质细胞。目前认为它们是牙髓干细胞,有高度增殖、自我更新的能力和多向分化的潜能。该群细胞平时位于血管旁的微环境中,保持未分化的静止状态,在牙髓损伤和修复过程中,可以自我复制并向成牙本质细胞、成纤维细胞等功能细胞分化,替代受损的细胞,维持局部组织稳态平衡。牙髓干细胞并没有非常显著的细胞表面标志物,STRO-1被认为是较好的表面标记。在体外诱导环境牙髓干细胞可向骨向分化、脂向分化。
-----------------------------------------------------------------------------------------
◇ 树突状细胞(dendritic cells),是近年来得到证实的牙髓中的细胞。此细胞见于整个牙髓,但主要分布在牙髓中央区的血管周围和牙髓的外周区如成牙本质细胞周围。此细胞常常有3个以上的胞质突起,长径可达50μm。树突状细胞表达HLA-DR抗原,在功能上属抗原呈递细胞,其特征是有树枝状胞质突起,细胞表面有II级组织相容性抗原,在诱导T细胞依赖性免疫中前关键作用。象抗原呈递巨噬细胞一样,他们吞噬蛋白抗原,然后呈递抗原的多肽片断和II级分子的组装体。T细胞可以识别这种组装体,这些片断与T细胞受体结合后使T细胞激活。因此该细胞是牙髓免疫防御系统中重要的组成部分。
-----------------------------------------------------------------------------------------
◇ 淋巴细胞,以往认为正常无炎症牙髓组织中无淋巴细胞。但研究证明,T淋巴细胞是正常牙髓中的一种重要的细胞。包括有CD4和CD8阳性细胞。它们是牙髓中主要免疫反应细胞。
牙髓中还有血管周细胞、血管内皮细胞和施万细胞等。
【纤维】
牙髓间质内主要是胶原纤维和嗜银纤维,而弹力纤维仅存在于较大的血管壁上。牙髓中的胶原纤维主要由I型和Ⅲ型纤维以55:45的比例所组成,纤维交织成网状。随着年龄的增加,胶原纤维的量逐渐增加,但其构成比则基本保持不变。嗜银纤维即网状纤维,为纤细的纤维,主要构成也是Ⅲ型胶原蛋白,分布于牙髓细胞之间。嗜银纤维只有在应用银染色时才能显示。其着色机制被认为与Ⅲ型胶原表面的蛋白多糖成分有关。牙髓也有少量Ⅴ型和VI型胶原。在牙本质形成的早期,在牙髓边缘聚集有粗大的科尔夫纤维束。
【基质】
牙髓中的基质是无定型的胶样物质,富含阴离子多糖,与牙髓组织含水的性质有关。牙髓中主要的蛋白多糖包括透明质酸、磷酸皮肤素、磷酸肝素和磷酸软骨素。牙萌出时,牙髓的蛋白多糖含量降低50%。牙本质形成活跃时,磷酸软骨素是是主要的蛋白多糖。牙萌出时,透明质酸和硫酸皮肤素增多,磷酸软骨素明显减少。蛋白多糖的功能是支持细胞、充盈组织、调节各种细胞的相互作用;影响细胞的黏附、活动性、生长和分化。基质也是一个分子筛,可阻挡大分子蛋白质通过。细胞代谢产物、营养物质和水分可通过细胞和血管间的基质。
【血管】
 牙髓内血管丰富。血管来自颌骨的牙槽动脉分支,它们经根尖孔进人牙髓,改称为牙髓动脉,沿牙髓中轴前进,沿途分出小支,最后在成牙本质细胞层下方形成一稠密的毛细血管丛,冠部尤其是髓角处毛细血管网密集。毛细血管后静脉汇成牙髓静脉与牙髓动脉伴行,出根尖孔转为牙槽静脉。
牙髓内血管丰富。血管来自颌骨的牙槽动脉分支,它们经根尖孔进人牙髓,改称为牙髓动脉,沿牙髓中轴前进,沿途分出小支,最后在成牙本质细胞层下方形成一稠密的毛细血管丛,冠部尤其是髓角处毛细血管网密集。毛细血管后静脉汇成牙髓静脉与牙髓动脉伴行,出根尖孔转为牙槽静脉。
牙髓和牙周膜的血管除通过根尖孔交通外,尚可通过一些副根管相通。因此,当牙髓或牙周组织发生炎症时,也可沿此通道相互扩散。牙髓中最大的动脉直径50~100μm,与身体其他部位的小动脉相似。这些动脉内膜为扁平或立方的内皮细胞,中膜有1~3层平滑肌细胞,外膜有少量胶原纤维。小动脉直径为20~30μm,中膜有1~2层平滑肌细胞,外膜与细胞间质的纤维相融合。末梢动脉位于牙髓边缘,直径为10~15μm,在内皮细胞外仅有一层平滑肌细胞围绕。毛细血管直径为8~10μm,仅见一层内皮细胞,甚至于在部分区域只见一层胞质膜。近成牙本质细胞的毛细血管部分为有孔毛细血管,孔的开口处内皮细胞的连续性中断,仅有一层基底膜覆盖。此孔能为邻近的成牙本质细胞液体和代谢物质的快速转运通道。牙髓的静脉较同名动脉为大,最大的静脉直径为100~150μm,管壁较动脉不规则,与身体其他部位同样大小的静脉相比,管壁相对较薄,内皮细胞扁,胞质不突向腔内,中膜为1~2层平滑肌细胞,在较小的静脉该层不完整或缺乏,无外膜。牙髓中可见动、静脉吻合,是动、静脉不经过毛细血管直接交流的通道,被认为在牙髓炎症和损伤时调节血液循环的重要结构,可减轻炎症或损伤时的组织压力。
【淋巴管】
牙髓中淋巴管常与血管伴行。淋巴毛细管起于牙髓表面,汇合成较大的小淋巴管,经髓核,穿过根尖孔与牙龈、牙周膜的淋巴管丛吻合。前牙的淋巴液引流入颏下淋巴结,后牙的则引流入下颌下和颈深部淋巴结。牙髓的淋巴管在光镜下不易与毛细血管区别。
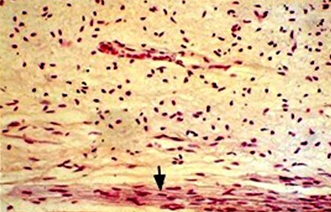
【神经】
牙髓内的神经很丰富(人前磨牙经根管口进入牙髓的神经轴突可达2000条或更多)。感觉神经和节后交感神经分别来自三叉神经和颈上神经节。伴同血管自根尖孔进入牙髓,并逐渐分成很多更细的分支。进入牙髓的两种感觉神经为有髓A-β、A-δ纤维和无髓C纤维。有髓纤维与各种伤害的感受有关;无髓纤维的传入部分与伤害感受有关,传出部分为节后交感神经纤维。有的神经纤维在根髓内即发出分支,有些在冠髓及髓角处、甚至在牙本质小管内才发出分支。在髓室内神经纤维分散呈放射状,近多细胞层处形成神经网,称为神经壁层(parietal layer of nerves)或Raschkow丛。自此层神经轴突通过多细胞层、无细胞层和成牙本质细胞层,止于牙髓牙本质交界处的成牙本质细胞突起之间或牙本质小管内。神经末梢呈圆或椭圆形膨大,与成牙本质细胞紧密相接。牙髓内的神经大多数是有髓神经,传导痛觉,少数为无髓神经,系交感神经,可调节血管的收缩和舒张。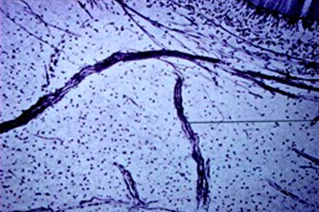
多数冠髓和冠髓内的神经末梢与血管分布相关,含有致密核的空泡,儿茶酚氨阳性染色,提示这些神经末梢可调节牙髓血流。在成牙本质细胞区,神经末梢含有大量含致密核的小泡,这些神经末梢与毛细血管关系密切,可能有调节成牙本质细胞对伤害的反应或成牙本质细胞的代谢的作用。
神经组织化学和免疫组织化学研究显示,牙髓神经中含有降钙素基因相关肽(CGRP)、enkephalin、神经肽Y、血管肠肽(VIP)、P物质、somatostatin、血清素、乙酰胆碱、去甲肾上腺素等神经递质。在组织损伤、补体激活、抗原抗体反应和逆行刺激下牙槽神经时,可使它们释放,引起血管的变化。CGRP和P物质还可引起痛觉过敏和启动创伤愈合。

在牙发育完成,即根尖孔形成以后,随着年龄的增长和牙受到外界的生理或病理性刺激,继发性牙本质和(或)修复性牙本质等不断形成,可使髓腔逐渐缩小。同时,牙髓组织中的细胞成分逐渐减少。成牙本质细胞由高柱状变为矮柱状或扁平,部分成牙本质细胞凋亡,剩余的成牙本质细胞对刺激的反应缓慢。成纤维细胞数量减少,同时伴纤维的数量和大小的增加。血管中可出现机体其他部位出现的胆固醇沉积,可使管壁黏附性增加并引起局部炎症反应。牙髓活力降低,出现退行性改变。
虽然牙髓位于髓腔内,但却凭借成牙本质细胞突起与外界有着密切的联系。任何物理和化学的刺激加到牙本质表面时,与该部位相应的牙髓组织必然发生反应。若所受刺激是慢性的、较弱的,则可引起修复性牙本质形成,并可部分造成牙髓组织的各类退行性变;若所受的刺激强烈,则可发生炎症反应。当牙髓发生炎症时,由于牙髓内的血管管壁薄,易于扩张、充血及渗出,使髓腔内的压力增大,而四周又为坚硬的牙本质壁所包围,无法相应扩张以减轻压力,牙髓神经末梢受压而产生剧烈疼痛。
牙髓内的神经在受到外界刺激后,常反应为痛觉,而不能区分冷、热、压力及化学变化等不同感受。这可能是因为牙髓缺乏对这些刺激的感受器。此外,牙髓神经还缺乏定位能力,故牙髓炎患者往往不能准确指出痛牙的部位。
牙髓是结缔组织,有修复再生的能力。但由于牙髓的解剖条件所限,其修复再生能力是有限的。当牙髓受到非感染性的较轻损伤时,修复一般是良好的。对于新鲜暴露的牙髓,经适当的临床治疗后,牙髓内的未分化间叶细胞可分化为成牙本质样细胞,形成牙本质桥。而当牙髓由于感染而发生炎症时,则完全的修复性再生是困难的。这对临床牙髓病的治疗具有参考价值。
-----------------------------------------------------------------------------------------

