
细胞信号转导系统(cell signaling system)由细胞信号、接受信号的受体或类似于受体的物质、细胞内信号转导通路及细胞内的效应器组成。
一、细胞信号转导过程
1. 细胞信号的种类
(1)化学信号
是生物体内最主要的信号,
可以识别并特异地与细胞受体结合的、具有生物活性的化学信号物质被称为配体(ligand),包括细胞间化学信号和细胞内化学信号。
细胞间化学信号:
①可溶性的化学分子,如激素、神经递质和神经肽、细胞生长因子和细胞因子、细胞的代谢产物(如ATP、活性氧等)以及药物和毒物包括细菌毒素等;
②气体分子;
③细胞外基质成分和与质膜结合的分子(如细胞黏附分子等)。
细胞内化学信号:
①环核苷酸如cAMP和cGMP等;
②脂质信使分子如甘油二酯(DAG)和三磷酸肌醇(IP3)等;
③气体信使分子如一氧化氮( NO)和一氧化碳(CO);
④离子信使分子如Ca2+和H+等。
(2)物理信号
包括各种射线、光信号、电信号、机械信号(摩擦力、压力、牵张力及切应力等)以及冷热刺激等。
已证明物理信号能激活细胞内的信号转导通路,如视网膜细胞中的光受体,可以感受光信号并弓|起相应的细胞信号系统激活。
(3)生物大分子的结构信号
这类信号常常包含在决定生物大分子(蛋白质、多糖类、核酸)三维结构的序列中。
以结构信号为基础的分子识别在细胞信号转导中具有重要的作用:
①决定细胞间识别和黏附;
②决定信号分子与受体的识别和结合;
③决定细胞信号转导通路中信号转导分子的连接及信号复合物的形成。
2. 细胞信号的接受和转导
细胞信号由受体或类似于受体的物质接受,然后将信息转发到细胞内,启动细胞信号转导过程。
(1)细胞受体
受体是细胞表面或亚细胞组分中的一种分子,可以识别并特异地与配体结合,从而激活或启动一系列生物化学反应,最后导致该信号物质特定的生物学效应。
根据分布部位可分为膜受体与细胞内受体。膜受体占受体的大多数,细胞内受体主要是核受体超家族。
膜受体
膜受体一般为跨膜糖蛋白,具有膜外区、跨膜区和细胞内区。
据它们的分子结构不同,可分为:
G蛋白耦联受体( G protein coupled receptor , GPCR)家族;
酪氨酸蛋白激酶型受体( tyrosine-specific protein kinase receptor , RPTK)家族;
酪氨酸蛋白激酶关联受体( tyrosine protein kinase-linked receptor , TPKR)家族;
丝/苏氨酸蛋白激酶( PSTK)型受体家族;
死亡受体家族(如TNFR、Fas等) ;
离子通道型受体家族( N-乙酰胆碱受体、N-甲基-D-门冬氨酸受体、环核苷酸受体、三磷酸肌醇受体、ryanodine受体等) ;
细胞黏附分子(如钙黏素、整合素等)等。
核受体
核受体本质上为一类配体依赖的转录调节因子。
受体与配体结合后,主要通过调节靶基因的表达产生生物学效应。
糖皮质激素受体( glucocorticoid receptor , GR) ;
性激素受体( SHR) ;
甲状腺激素受体( thyroid hormone receptor ,TR);
1 ,25-(OH)2D3受体( VDR) ;
维A酸受体( RAR) ;
代谢性受体;
小分子气体受体等。
另外,目前发现的绝大部分孤儿受体( orphan receptor)亦属于核受体。
(2)细胞信号转导的基本过程
细胞信号转导过程是将细胞信号通过受体或类似物质将信号导入细胞内并引起细胞内一系列信号转导蛋白的构象、活性或功能变化,从而实现调控细胞结构和功能的作用。
细胞信号转导的过程十分复杂,而且存在广泛的细胞通路间的交叉调控。
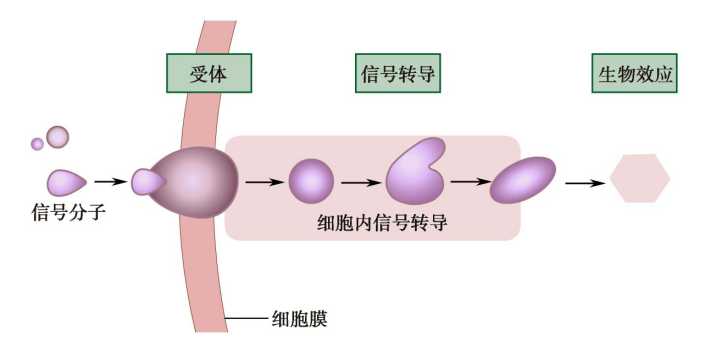
图10-1 细胞信号转导基本过程
3. 常见的细胞信号转导通路
(1)G蛋白耦联受体介导的信号转导途径
腺苷酸环化酶信号转导通路
在腺苷酸环化酶信号转导通路中存在着两种作用相反的G蛋白:刺激性G蛋白Gs与抑制性G蛋白Gi。它们通过增加或抑制腺苷酸环化酶活性来调节细胞内cAMP浓度,进而影响细胞的功能。
β肾上腺素受体、胰高血糖素受体等激活后与Gs偶联,活化的Gsα增加腺苷酸环化酶的活性,促进cAMP生成。而α2肾上腺素受体、M2胆碱受体及血管紧张素Ⅱ(angiotensin II,AngⅡ)受体等激活则与Gi偶联,抑制cAMP的生成。cAMP可激活蛋白激酶A(protein kinase A,PKA),引起多种靶蛋白磷酸化,调节其功能。
例如,肾上腺素引起肝细胞内cAMP增加,通过PKA促进磷酸化酶激酶活化,增加糖原分解。心肌β肾上腺素受体兴奋引起的cAMP增加经PKA促进心肌钙转运,提高心肌收缩力。进入核内的PKA可磷酸化转录因子CRE结合蛋白(cAMP response element binding protein,CREB),使其与靶基因DNA调控区的cAMP应答元件(cAMP response element,CRE)结合,激活靶基因转录(图 10-1)。
磷脂酶C信号转导通路
α1肾上腺素受体、内皮素受体和AngⅡ受体等激活可与Gaq结合,激活细胞膜磷脂酶C(phospholipase C,PLC)β亚型。PLCβ催化磷脂酰肌醇二磷酸(phosphatidylinositol 4,5-diphosphate,PIP2)水解,生成三磷酸肌醇(1,4,5-inositol triphosphate, IP3)和甘油二酯(1,2-diacylglycerol,DAG)。IP3促进肌浆网或内质网储存的Ca2+释放,Ca2+可作为第二信使启动多种细胞反应。例如,激发胰岛β细胞的胞吐作用,促进胰岛素释放;与心肌和骨骼肌的肌钙蛋白结合,触发肌肉收缩。
Ca2+还可与钙调素(calmodulin, CaM)结合,激活Ca2+/CaM依赖性蛋白激酶,经磷酸化靶蛋白或转录因子产生生物学作用。DAG与Ca2+协调促进蛋白激酶C (protein kinase C, PKC)活化。激活的PKC催化靶蛋白磷酸化,改变其生物活性。例如,PKC促进细胞膜Na+-H+交换蛋白磷酸化,增加H+外流;促进表皮生长因子受体磷酸化,参与受体的下调。激活的PKC也可通过磷酸化转录因子AP-1和NF-κB等,诱导c-fos、c-jun的表达,促进靶基因转录和细胞增殖肥大(图10-1)。
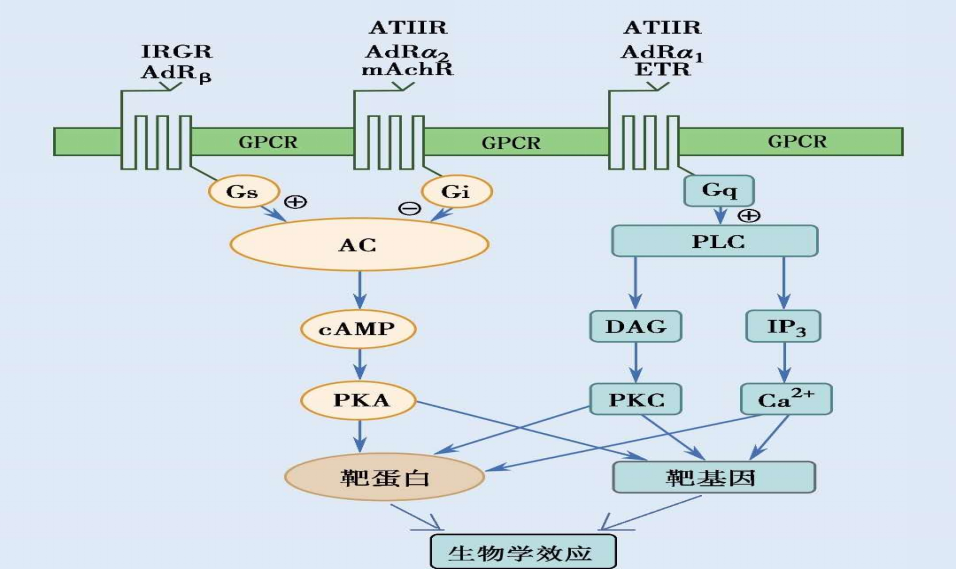
图10-2 GCPR介导的细胞信号转导途径示意图
(2)受体酪氨酸蛋白激酶介导的信号转导途径
受体酪氨酸蛋白激酶是含有内在酶结构的受体超家族,其共同的结构特征是单次跨膜受体,配体以生长因子为代表。
丝裂原活化蛋白激酶(mitogen activited protein kinase,MAPK)家族是一个高度保守的丝/苏氨激酶大家族,至少包括4个亚家族:ERK1/2、p38、JNK和ERK5等,在与细胞生长、分化和凋亡等密切相关的信号转导途径中发挥重要的调节作用。
MAPK家族可由多种方式激活,其中研究较多的是Ras-Raf-MEK-ERK途径。EGF和PDGF等生长因子与其受体结合并引起受体酪氨酸蛋白激酶激活后,细胞内含SH2区的生长因子受体连接蛋白(growth factor receptor-bound protein 2,Grb2)识别受体上磷酸化的酪氨酸并转移至质膜,将胞浆中具有鸟苷酸交换因子活性的Sos吸引至细胞膜,Sos促进无活性Ras所结合的GDP为GTP所置换,导致Ras活化。激活的Ras活化丝/苏氨酸激酶Raf(又称MAPK kinase kinase, MAPKKK),进而激活MEK(MAPK kinase,MAPKK),最终导致细胞外信号调节激酶(extracellular signal regulated kinase,ERK)1/2激活。激活的ERK1/2可促进胞浆靶蛋白磷酸化或调节其他蛋白激酶的活性,如激活磷脂酶A2;激活调节蛋白质翻译的激酶等。ERK入核可使多种转录因子磷酸化,如促进血清反应因子(serum response factor, SRF)磷酸化,使其与含有血清反应元件(serum response element, SRE)的靶基因启动子相结合,增强转录活性(图10-2)。

图10-3 RPTK介导的细胞信号转导途径示意图
(3)非受体酪氨酸蛋白激酶介导的信号转导途径
非受体酪氨酸蛋白激酶属于与胞浆中蛋白激酶偶联的受体家族,是单次跨膜受体,当其与配体结合并激活后,可和胞浆内含酪氨酸蛋白激酶的分子相结合。
配体主要是激素和细胞因子。
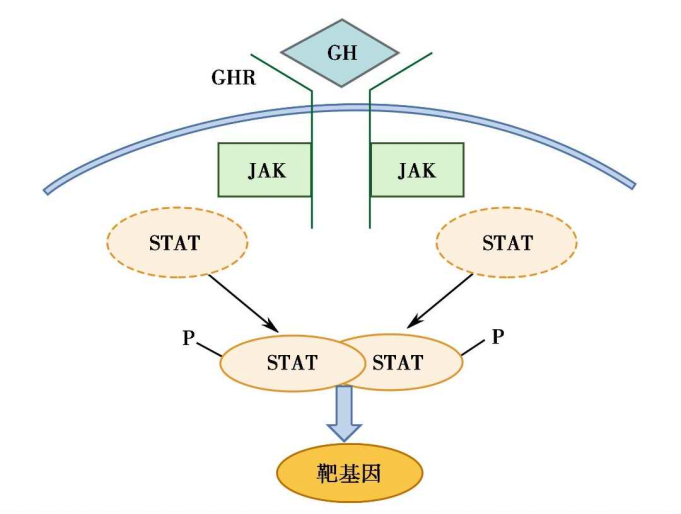
图10-4 GHR介导的细胞信号转导途径示意图
(4)核受体介导的信号转导途径
按核受体的结构与功能可将其分为:
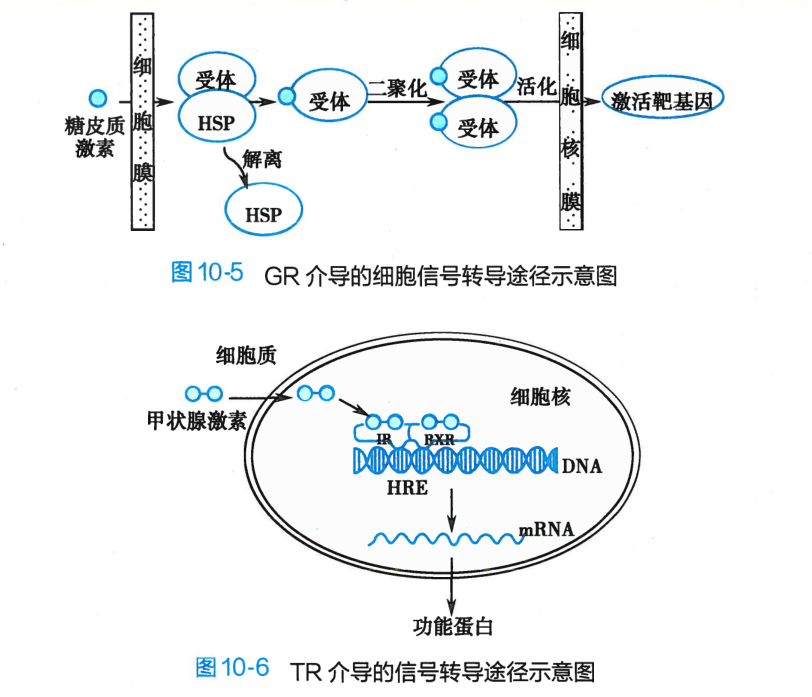
①类固醇激素受体家族,包括糖皮质激素、盐皮质激素、性激素受体等。类固醇激素受体家族位于胞浆(除雌激素受体位于核内),未与配体结合前与热休克蛋白(heat shock protein,HSP)结合存在,处于非活化状态。配体与受体的结合使HSP与受体解离,暴露DNA结合区。激活的受体二聚化并转移入核,与DNA上的激素反应元件(hormone response element, HRE)相结合或与其他转录因子相互作用,增强或抑制靶基因转录(图10-5);
②甲状腺素受体家族,包括甲状腺素、维生素D和维甲酸受体等。此类受体位于核内,多以同源或异源二聚体的形式与DNA或其他蛋白质结合,配体入核与受体结合后,激活受体并经HRE调节基因转录(图10-6)。
此外,还有代谢性核受体家族等。
二、细胞信号转导的调节
1. 信号调节
根据配体引发细胞反应的结果不同,将其分为两大类:激动剂与拮抗剂。
前者与受体结合可激活受体的内在活性;后者与受体结合可阻抑激动剂与受体结合,从而抑制激动剂的作用。
配体一般通过两种方式控制信号转导蛋白的活性:
(1)配体与信号蛋白结合直接改变信号蛋白活性
如细胞内信使分子cAMP与二酰甘油(DAG)能分别激活蛋白激酶A( PKA)和蛋白激酶C( PKC)。
(2)配体通过激活受体型蛋白激酶控制信号转导
如细胞外信号(如胰岛素)可激动酪氨酸蛋白激酶型受体-胰岛素受体,通过激活多条信号转导通路控制糖、蛋白质代谢及细胞增殖等功能。
2. 受体调节
(1)受体数量的调节
受体下调:当体内配体持续升高时,配体受体复合物可被细胞内化,内化后配体及部分受体被降解,部分受体返回胞膜重新利用,可致自身受体数量减少。受体下调可引起该受体介导的信号转导抑制
受体上调:持续高浓度的配体与受体结合,除可引起自身受体下调外,还可引起其他受体明显增多。受体上调可引起该受体介导的信号转导加强
(2)受体亲和力的调节
受体增敏:受体对配体刺激的反应增强,可引起该受体介导的信号转导加强
受体减敏:受体对配体刺激的反应衰退,可引起该受体介导的信号转导抑制
在通常情况下,受体上调与受体增敏相联系,受体下调与受体减敏相关联。
3. 受体后调节
(1)通过可逆磷酸化快速调节靶蛋白的活性
信号转导通路对靶蛋白调节的最重要方式是可逆性的磷酸化调节。
多种信号转导通路中激活的蛋白激酶(如PKA、PKB、PKC、MAPK家族中的成员等)和磷酸酶能通过对各种效应蛋白(如调节代谢的酶、离子通道、离子泵、运输蛋白、骨架蛋白等)及转录因子(如NF-κB、AP-1等)进行可逆的磷酸化修饰,快速调节它们的活性和功能,产生相应的生物学效应。
(2)通过调控基因表达产生较为缓慢的生物效应
胞外信号调节基因转录有两种方式:
一是胞外信号启动细胞的信号转导,在信号通路中激活的蛋白激酶首先磷酸化细胞中现存的转录因子,使其激
活并转入胞核,启动相应基因的转录过程;
二是某些信号可直接进人细胞(如甾体激素),与核受体结合,调节靶基因的表达而产生较为缓慢的生物学效应。

