
8. 植物的生长生理
视频:chapter 8 physiology of plant growth.mp4(下载附件 1.01 GB)
课件:chapter 8 Physiology of plant growth.ppt(下载附件 9.5 MB)
中文课件:
提要:植物生长(plant growth)是指植物在体积和重量上的不可逆增加,是由于原生质体的增加和细胞壁的伸展,引起细胞分裂、生长和分化,以及植物体新器官的形成和分化。种子的萌发就是一个生长过程。植物从种子萌发到幼苗形成,经历了从异养到自养的转变。幼苗的形成标志着自养体系的建成,从此植物进入了营养体迅速发展时期。这一时期的主要特征是营养器官(根、茎、叶)的旺盛生长,这是以旺盛的代谢活动为物质基础的。在此基础上,植物体不断地成长壮大,到一定的时期就开花结实,形成新的种子。本章介绍细胞分裂、分化生理,种子的萌发,根、茎、叶等营养器官生长的规律以及与植物生长密切相关的植物组织培养、植物运动机理等方面的知识。
8.1种子的萌发
8.1.1 种子萌发的概念
种子是由受精胚珠发育而来的种子植物所特有的延存器官。植物个体的生命周期是从受精卵分裂形成胚开始的,但人们习惯上还是以种子萌发作为个体发育的起点。所谓种子萌发(seed germination)是指种子从吸水到胚根突破种皮期间所发生的一系列生理生化变化过程。风干种子的生理活动极为微弱,处于休眠状态。在有足够的水分、适宜的温度和正常的空气中,种子开始萌发。从形态上看,种子萌发是具有生活力的种子吸水后,胚生长突破种皮并形成幼苗的过程。通常以胚根突破种皮作为萌发的标志。从生理上看,萌发是无休眠或已解除休眠的种子吸水后由相对静止状态转为生理活动状态,呼吸作用增强,贮藏物质被分解并转化为可供胚利用的有机物,使胚生长的过程。
种子萌发过程大致可分为以下步骤:种子吸水萌动;内部的物质与能量转化;胚根突破种皮形成幼苗。但是,这些过程不是彼此孤立,而是相互联系的。
8.1.2 种子的寿命和活力
8.1.2.1 种子的寿命
种子的寿命(seed longevity)是指种子从成熟到丧失发芽能力所经历的时间。根据植物种类和所处条件的不同,在自然条件下,种子的寿命可以由几小时到很多年。寿命极短的种子如柳树种子,成熟后只在12 h内有发芽能力。杨树种子的寿命一般不超过几个星期。大多数栽培作物如水稻、小麦、大麦、大豆、菜豆等的种子寿命1~3 年。少数种子寿命较长的,如蚕豆、绿豆、豇豆、紫云英等能达5~11年。种子寿命长的甚至可达百年以上。北京植物园曾对从泥炭土层中挖出的沉睡千年的莲子进行催芽,莲子竟仍能发芽和正常开花结果。远在更新世(距今1万年前)时期埋入北极冻土层淤泥中的北极羽扁豆种子,挖出后可在实验室里迅速萌发,这些都是长命种子。
种子寿命的长短主要是由遗传基因决定的,但也受环境因素、贮藏条件等的影响。根据植物种子保存期的特点,将种子分为正常性种子(orthodox seed )和顽拗性种子(recalcitrant seed )。正常性种子耐脱水性很强,通常在干燥低温状态下可长期贮藏而不丧失活力,大多数植物种子属于这一类。而顽拗性种子是不耐脱水干燥,也不耐零下低温贮藏。产生顽拗性种子的植物主要有两大类:(1)原产于热带或亚热带地区的许多果树,如椰子、荔枝、龙眼、芒果等;(2)一些水生草本植物,如水浮莲、茭白、菱等。
关于正常性种子耐脱水和顽拗性种子不耐脱水的机理,现在认为可能与LEA基因的表达有关。LEA基因在种子发育晚期表达,其产物被称为胚胎发育晚期丰富蛋白(lateembryogenesis abundant protein ,LEA)。LEA的特点是具有很高的亲水性和热稳定性,并可被ABA和水分胁迫等因子诱导合成。LEA在种子成熟脱水过程中起到保护细胞免受伤害的作用。LEA蛋白在成熟的正常性种子中含量很高,如在棉花的成熟种子中,其约占贮藏性蛋白的30% 。对于顽拗性种子,由于这些植物大多生长在温湿地区或水域中,随时具备适宜萌发的环境,一旦种子成熟即可萌发,毋须经过脱水阶段,因而体内LEA蛋白积累不多,表现出对脱水的敏感性。
8.1.2.2种子的生活力与活力
种子生活力(seed viability)是指种子能够萌发的潜在能力或种胚具有的生命力。一般来说,种子的生活力就是指种子的发芽力。没有生命力的种子是不能萌发的。为了准确、全面地地评价种子萌发质量,人们又引入了种子活力的概念。所谓种子活力(seed vigor),是指种子在田间状态(即非理想状态)下迅速而整齐地萌发并形成健壮幼苗的能力。在一般情况下,种子的活力水平随着种子的发育而上升,至生理成熟期达到高峰。成熟后的种子,因品种特性和贮藏条件不同,其生理机能以不同的速度发生恶化,活力水平也就逐渐下降。种子活力的高低,可直接影响农作物生产。在播种时选用高活力的种子有利于形成健壮的幼苗,从而提高作物的抗逆能力和增产潜力。
8.1.3 影响种子萌发的外界条件
有生活力并已破除休眠的种子要萌发,还需有适宜的外界环境条件。影响种子萌发的外界条件主要有水分、温度、氧气,有些种子的萌发还受光的影响。
8.1.3.1水分
吸水是种子萌发的第一步。种子只有在吸收了足够的水以后,各种与萌发有关的生理生化作用才能逐渐开始。这是因为水分可使种皮变软,氧气容易透过种皮,增加胚的呼吸,同时也使胚易于突破种皮;水分可使原生质由凝胶状态转化为溶胶状态,使酶活性提高,为呼吸、物质转化、运输等一系列代谢活动提供基本条件;促进不溶性的大分子化合物转化为可溶性的低分子化合物,供胚萌发生长之用;使种子内储藏的植物激素由束缚型转化为游离型,以调节胚的生长。因此,充足的水分是种子萌发的首要条件。
在萌发过程中,种子吸水的程度和速率与种子成分、温度以及环境中水分的有效性有关。一般淀粉和油料种子吸水达风干重的30~70%即可发芽,蛋白质含量高的种子吸水要达风干重的110%以上才能发芽,这是因为蛋白质有较大的亲水性。表8.1列举了几种主要作物种子萌发时的吸水量。
表8.1 各种主要作物种子萌发时的最低吸水量(占风干重的百分率)
| 作物种类 吸水率(%) | 作物种类 吸水率(%) |
| 水稻 35 小麦 60 玉米 40 油菜 48 | 棉花 60 豌豆 186 大豆 120 蚕豆 157 |
种子吸水速率不仅与种子内贮藏物质的种类有关,还受土壤含水量、土壤溶液浓度以及环境温度的影响。一般情况下,土壤含水量充足,土壤溶液浓度较低,环境温度较高,均能促进种子吸水。
8.1.3.2 温度
种子萌发过程中的一系列生理生化过程,是在一系列酶的催化下完成的,而酶促反应与温度密切相关,因此温度也是影响种子萌发的一个重要的外界条件。温度对种子萌发的影响有三基点,最低、最高和最适温度。最低和最高温度是种子萌发的极限温度,低于最低温度或超过最高温度,种子都不能萌发。最适温度是指在最短的时间内种子萌发率最高的温度。常见作物种子萌发的温度范围见表8.2。
表8.2 几种作物种子萌发的温度三基点
| 作物种类 | 最低温度/0C | 最适温度/0C | 最高温度/0C |
| 玉米、高粱类 大、小麦类 水稻 棉花 大豆 花生 黄瓜 番茄 | 8 ~ 10 3 ~ 5 10 ~ 12 10 ~ 12 6 ~ 8 12 ~ 15 15 ~ 18 15 | 32 ~ 35 20 ~ 28 30 ~ 37 25 ~ 32 25 ~ 30 25 ~ 37 31 ~ 37 25 ~ 30 | 40 ~ 45 30 ~ 40 40 ~ 42 38 ~ 40 39 ~ 40 41 ~ 46 38 ~ 40 35 |
各种种子所要求的适宜的萌发温度,一般与其原产地的生态条件有密切关系。萌发的最适温度,尽管是生长最快的温度,但由于种子消耗的有机养分较多,往往使幼苗生长很快但并不健壮,经不起不良环境侵袭。所以生产上常采用比萌发最适温度稍低的协调最适温度,即指生长快而又健壮的温度。掌握萌发的最低温度和最高温度,在生产上是决定不同播种期的主要依据。适宜的播种期一般以稍高于最低温度为宜,如棉花播种期一般以表土五厘米土温稳定在12℃为宜。为了提早播种,早稻可进行薄膜育秧,其他植物可利用温室、大棚、温床、阳畦、风障等设施育苗。
变温处理(通常是低温下16h,高温下8 h,其变温幅度大于10℃)有利于种子的萌发,而且还可提高幼苗的抗寒力。自然界中的种子大都是在变温的情况下萌发的。
8.1.3.3 氧气
种子萌发是活跃的生长过程,要进行旺盛的物质代谢,包括合成原本不存在于干种子中的酶(如α- 淀粉酶),贮藏在胚乳或子叶中的有机物被分解并运输到胚根、胚芽进行再合成等。这些反应过程所需的能量主要由有氧呼吸提供,因此,种子萌发过程中必需要有充足的氧气供应。如果种子萌发期间供氧不足则导致无氧呼吸,一方面贮藏物质消耗过多过快,另一方面无氧呼吸产生酒精引起中毒。在农业生产上,实行土壤深耕、平整土地、改良土壤、中耕松土等措施,目的之一就是为了增加土壤中的氧气,改善土壤的通气条件。
种子的种类不同,萌发时对氧的要求也不同。一般种子正常萌发要求空气中氧浓度在10%以上,当氧浓度在5 %以下时,大部分种子不能萌发。含脂肪较多的种子,如花生、大豆和棉花等萌发时,比淀粉种子要求更多的氧气。水稻对缺氧的忍耐能力较强,其种子在淹水的情况下能依靠无氧呼吸来萌发。但缺氧时,水稻萌发后只长胚芽鞘,不长根,幼苗生长也十分细弱。若无氧呼吸进行太久,会消耗较多的有机物而释放能量少,还会积累酒精使种子中毒等。胚芽鞘的生长,只有细胞生长而没有细胞分裂,仅靠无氧呼吸产生的能量已可发生;而胚根和胚芽的生长,既有细胞生长又有细胞分裂,只有依赖有氧呼吸才能满足它们对能量和物质的需求。因此,在水稻育秧时,要注意秧田排水,以保证供氧充足。
8.1.3.4光
大多数作物的种子萌发时对光照不敏感,只要水分、氧气和温度条件满足了就能萌发。
但有些植物,如莴苣、烟草、紫苏、桦木和许多杂草的种子,需要光照才能萌发,这类种子称为需光种子。而另一类植物,如韭菜、苋菜、番茄、茄子、瓜类等的种子,只能在暗处萌发,光照会抑制萌发过程,这类种子称为需暗种子。光对种子萌发的影响是通过光敏色素(见7 光形态建成)实现的。
8.1.4种子萌发时的生理生化变化
具有生活力并已解除休眠的种子在满足水分、氧气、温度以及光照等条件后,就进入种子萌发的过程。种子萌发包括胚的萌动与生长和胚乳或子叶中贮藏产物的动员。种子的贮藏物质需要经酶的分解后才能以可利用的方式动员到胚中去,以支持胚的迅速生长。种子萌发过程中的生理生化变化主要包括:种子的吸水,呼吸作用的变化,酶的活化和合成及种子中贮藏物质的动员等过程。
8.1.4.1 种子的吸水
种子萌发过程中的吸水可分为三个阶段(图8.1)。第一阶段,种子吸水迅速,这一阶段的吸水是一个物理过程,称吸胀作用(imbibition)。由于这个阶段的吸水主要依赖种子的衬质势,因此不论种子是否休眠,是否具有生命力,均能进行。第二阶段是种子吸水停滞期。经过第一阶段的快速吸水,原生质的水合程度趋向饱和;细胞膨压增加,阻碍了细胞的进一步吸水。活种子在这一阶段的代谢活动却非常旺盛,细胞利用已吸收的水进行各种代谢活动。第三阶段是生长吸水阶段。由于胚的迅速生长,胚根突破种皮,有氧呼吸加强,种子重新大量吸水,这时的吸水是与代谢作用相关的渗透性吸水。对于休眠或死亡种子却只停留在第二阶段的状态。
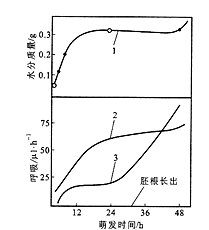
图8.1豌豆种子萌发时吸水和呼吸的变化
1. 种子吸水过程的变化;2. CO2的变化;3. O2吸收的变化
8.1.4.2 呼吸作用的变化
种子萌发过程中呼吸作用的变化与吸水过程相似,也分为三个阶段(图8.1)。种子吸水的第一阶段,呼吸作用也迅速增加,这主要是由已经存在于干种子中并在吸水后活化的呼吸酶及线粒体系统完成的。在吸水的停滞期,呼吸作用也停滞在一定水平,一方面是因为干种子中已有呼吸酶及线粒体系统已经活化,而新的呼吸酶及线粒体还没有大量形成;另一方面,此时胚根还没有突破种皮,氧气的供应也受到一定限制。吸水的第三阶段,呼吸作用又迅速增加,因为胚根突破种皮后,氧气供应得到改善,而且此时生长的胚轴细胞合成新的线粒体和呼吸酶系统。
由图8.1可见在种子吸水的第一和第二阶段, R.Q>1,到第三阶段,胚根长出,种子鲜重持续增加,R.Q<1。这说明种子萌发的初期主要是无氧呼吸,而随后进行的是有氧呼吸8.1.4.3 酶的变化
种子萌发时,由休眠状态转入生理活动活跃状态,需进行酶的活化和合成过程。种子的贮藏物质也需要经过分解后,才能以可利用的形式动员到胚中去,这样就需要一些分解酶类的迅速合成。种子萌发时, 这些酶类有多种不同的来源:一是在种子形成时产生,即已经存在于干燥种子中,在种子吸水后立即具有活性的,如β- 淀粉酶、磷酸酯酶、支链淀粉葡萄糖苷酶以及呼吸系统的酶和蛋白质合成系统中的酶;二是由贮藏mRNA在萌发过程翻译而成的,一般在吸水几小时后就有活性;第三类酶活性在吸水后较晚时期才出现,这类酶可能是在种子吸胀后,由一些基因转录成新的mRNA合成新的酶,如α- 淀粉酶。
种子萌发所需的大多数酶需要在吸水后重新合成。酶重新合成所需的mRNA ,或已经存在于干燥种子中,或是种子吸水后由DNA转录而来。已经存在于干燥种子中的mRNA是在种子发育期间形成的,在种子萌发初期作为模板合成萌发所需要的蛋白质,人们把这类mRNA称为贮存mRNA(长命mRNA ,预先形成的mRNA等)。贮存mRNA对萌发早期几种水解酶的形成,以及胚根的发端可能起着重要作用。推测在种子形成时中合成的RNA有可能并不立即全部用于当时的蛋白质合成,其中一部分可用于编码种子萌发时蛋白质的合成。贮存mRNA可与细胞质中的蛋白质合成信息体(informosome)而保存在干燥种子中。除了贮存mRNA以外,在种子萌发过程中还有许多新的mRNA是由基因转录而来。
8.1.4.4有机物的转变
胚乳或子叶中贮藏有丰富的营养物质,主要为糖类、蛋白质和脂肪。不同植物种子中,这三类有机物的含量有很大差异。据此,将种子区分为淀粉种子(淀粉含量多)、油料种子(脂肪含量多)和豆类种子(蛋白质含量多)。这些贮藏物质在种子萌发时分解为简单的有机物,以可利用的形式运输到胚中,作为幼胚生长的营养物质。
1、碳水化合物的转变 种子萌发时,淀粉被淀粉酶、去分支酶(debranching enzyme)和麦芽糖酶等水解为葡萄糖。水解直链淀粉的淀粉酶包括α—淀粉酶和β—淀粉酶。α—淀粉酶是淀粉外切酶,它从直链淀粉上一次切下6个或12个葡萄糖分子,将淀粉水解为小分子的糊精;β—淀粉酶是淀粉外切酶,它从直链淀粉或糊精的末端葡萄糖起,每次切下一个麦芽糖分子。两者同时作用,可将直链淀粉完全水解为麦芽糖。麦芽糖在麦芽糖酶的作用下,可进一步分解为葡萄糖。支链淀粉除有α—1,4糖苷键外,还有分支处的α—1,6糖苷键。水解α—1,6糖苷键的酶是去分支酶,亦称R酶。淀粉在上述多种酶的作用下,逐渐被降解为分子量递减的各种糊精,最后被彻底水解为葡萄糖。不同分子量的淀粉和糊精与碘反应呈不同的颜色,可用于检查淀粉的水解程度。
除上述酶促水解外,淀粉也可以磷酸解(phosphorolysis)。在磷酸参与下,淀粉磷酸化酶把淀粉降解成葡萄糖-1-磷酸。淀粉磷酸化酶普遍存在于高等植物中。在禾谷类和豆类种子的萌发初期,淀粉的降解主要是依靠淀粉的磷酸化作用。到后期,淀粉的水解作用才成为淀粉降解的主要途径。淀粉降解的产物是以蔗糖形式从胚乳或子叶运输到生长中的胚芽和胚根中。
在种子萌发的早期,胚乳或子叶中的贮藏物质还不能用作萌发时,大多数干种子中含有一定量的蔗糖、棉子糖等寡糖。蔗糖的水解是在转化酶(亦称蔗糖酶)的作用下,水解为葡萄糖和果糖。
在种子萌发过程中,由于水解酶的作用,淀粉含量迅速下降,同时葡萄糖和其它单糖含量很快增加,以后又逐渐减少,这一方面是由于呼吸作用的消耗,另一方面是由于用作纤维素合成和蛋白质合成的碳骨架。在幼苗的生长过程中,不断形成新的细胞壁,所以纤维素的含量不断增加。
2、 脂肪的转变 大多数种子中贮藏的脂肪是甘油三酯。脂肪在脂肪酶的作用下,水解生成甘油和脂肪酸。由于脂肪酶的活性在酸性条件下较强,而脂肪水解所产生的脂肪酸可提高反应介质的酸性,所以脂肪酶具有自动催化的性质。
脂肪水解的产物甘油在酶的催化下,可氧化并磷酸化成磷酸丙糖(磷酸二羟丙酮、3-磷酸甘油醛),进一步通过糖的有氧氧化途径(三羧酸循环)而氧化分解为CO2和H2O,或逆糖酵解途径转变为葡萄糖、蔗糖等。
脂肪的另一种水解产物脂肪酸经过β—氧化形成乙酰辅酶A,然后经过乙醛酸循环途径形成琥珀酸,再进入线粒体进行TCA循环,产生苹果酸。苹果酸运到细胞质,被氧化为草酰乙酸,进一步羧化为磷酸烯酮式丙酮酸。再经过葡糖异生途径(gluconeogenic pathway),形成葡萄糖,最后转变为蔗糖,并转运至胚轴供生长之用。
3、蛋白质的转变
种子中的贮藏蛋白质积累在蛋白体中,禾谷类种子糊粉层中的蛋白体被称为糊粉粒。种子萌发时,贮藏蛋白质要从贮藏部位运到利用部位,以及由一种蛋白质转变为另一种蛋白质,都必须首先经过蛋白质水解过程。
蛋白质是在蛋白酶和肽酶的作用下水解为氨基酸的。首先蛋白质在蛋白酶的作用下分解为许多小肽,而后在肽酶的作用下完全水解为氨基酸。氨基酸在转氨酶的作用下,形成其他种类的氨基酸,使氨基酸种类增多,有利于新器官中蛋白质的合成。种子中的含氮化合物主要以酰胺(谷氨酰胺和天冬酰胺)的形式进行运输,运到新形成的器官中,重新合成蛋白质,供幼胚生长的需要。
种子中蛋白质水解产生的氨基酸,除了可以作为再合成蛋白质的原料外,也可以通过脱氨基作用,转变为有机酸和游离的氨(NH3)。有机酸可进入呼吸代谢途径,也可作为形成氨基酸的碳骨架。而氨对细胞有毒害作用,一般不会在细胞内积累,而是迅速转变为酰胺的方式贮存。以后在酶的作用下,酰胺又将NH3释放出来,供新的氨基酸合成之用。
综上所述,种子萌发时,胚乳中贮藏的碳水化合物、脂肪和蛋白质等大分子化合物,在各种水解酶的作用下,分解成各种简单的小分子化合物,从原来的不溶解状态变为可溶解状态。这些简单的有机物被运到正在生长的部位,经过合成作用形成新细胞的构成物质。种子萌发经历从异养到自养的过程。种子萌发初期只能动用种子内贮藏的物质,这就是异养。见光之后,开始了自养生活。当幼苗生出足够的叶片,进行旺盛的光合作用,制造充分的有机养料时,才进入完全自养生活。因此,种子内贮藏的养分越多,越有利于幼胚的生长。在农业生产上要选取粒大粒重的种子,就是这个道理。
种子萌发过程中,各种贮藏物质的分解和再利用可归纳为图8.2。
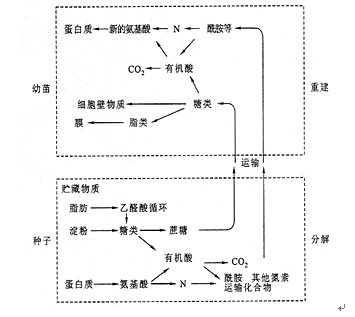
图8.2 种子萌发过程中贮藏物质的转变和再利用情况
8.1.5种子预处理与种子萌发的调节
种子是种子植物所特有的延存器官。播种活力高的种子是提高产量的必要条件。具有较高活力的种子,在田间状态下萌发迅速,能形成整齐度高而健壮的幼苗。而种子的活力水平随着种子的发育逐渐上升,至生理成熟期达到高峰。成熟后的种子处于脱水状态,并随着种子的采收、加工、贮藏、运输等,以不同速度发生劣变,活力水平又逐渐下降。在种子已发生劣变,活力有所下降的情况下,播种前进行种子的预处理,可以提高种子活力,改善其田间成苗状态, 这对干旱和盐碱地地区特别重要。如干旱和盐碱地地区小麦播种前,用过磷酸钙等预处理具有显著提高出苗率及增产作用。施用生理活性物质的形式有喷施、撒施、涂于种子表面(制成颗粒状或带状,即种子包皮)、通过有机溶剂渗入等,所施用的物质包括生长调节剂、矿质元素、杀虫剂、杀菌剂、杀鼠剂等。
对种子进行渗透调节(osmtic adjustment)处理可以缩短播种至出苗所需的时间,提高幼苗的整齐度。所谓渗透调节处理,一般是利用一定浓度的聚乙二醇(polyethylene glycol , PEG)溶液对种子进行处理。PEG是一种分子量较大的惰性物质,渗透调节处理中通常使用相对的分子质量为6000的PEG。种子在PEG溶液中吸水后开始萌动,进而引发细胞中的生理生化过程。可是由于PEG溶液具有一定的渗透势,因而可以控制水分进入细胞中的量,使萌发过程进行到一定程度后就停留在某一阶段而不能完成萌发的整个过程,这样所有种子的萌发最终都将停留在相同的阶段。一旦重新吸水后,所有种子都从相同阶段继续完成萌发过程,这样所产生的幼苗就具有较高的整齐度。近年的研究表明,渗透调节处理还可以促进萌发种子中RNA、蛋白质的合成,有利于种子中DNA损伤的修复等。
8.2 细胞的生长和分化
细胞的生长是植物体生长的基础。通常把细胞生长的过程分为三个时期:即分裂期、伸长期和分化期。通过细胞分裂增加细胞的数目,通过细胞伸长增加细胞的体积,通过细胞分化形成不同的组织和器官。
8.2.1 细胞分裂的生理
植物的分生组织细胞处在不断分裂的过程中。具有分裂能力的细胞,体积小、细胞质浓厚、无大液泡、细胞核大、细胞壁薄、合成代谢旺盛。植物分生组织的细胞生长到一定阶段就要发生有丝分裂,一分为二。新生的细胞长大后,再分裂为两个子细胞。通常把从母细胞一次分裂结束形成子细胞到下一次再分裂成两个子细胞之间的时期称为细胞周期(cell cycle)。细胞周期包括分裂间期(interphase)和分裂期(mitotic stage ,简称M期)。分裂期是指细胞的有丝分裂过程,也就是从染色体凝缩、分离到平均分配到两个子细胞为止,分裂后细胞内DNA减半。根据染色体形态指标可细分为前期、中期、后期和末期。分裂期以外的时间称为分裂间期,又分为三个时期:⑴ G1期(gap1),从有丝分裂完成到DNA复制之前的这段时间,此时细胞内进行RNA和蛋白质的大量合成,细胞体积也显著增大。⑵ S期(synthesis phase),DNA复制期,DNA和有关组蛋白在此时合成,完成染色体的复制,DNA的含量增加一倍。⑶ G2 期(gap2),从DNA复制完成到有丝分裂开始的这段时间,此时细胞继续进行RNA和蛋白质的合成,为细胞分裂做好准备(图8.3)。G2 期完成后,细胞进入分裂期。分裂形成的新细胞有的可以继续进行细胞周期的循环,另一些细胞可能转入G0期。G0期并非是细胞周期的一个阶段,G0期细胞是暂时离开细胞周期处于“静止”状态的细胞,在一定条件的诱导下可以重新进入细胞周期,进行增殖。
细胞周期的运转是十分有序的,沿着G1 →S→G2 →M的次序进行,这是细胞周期有关基因顺序表达的结果。与细胞分裂有关的cdc ( cell division cycle )基因的有序表达受细胞周期中一些控制点的调节控制,其中两个起决定作用的控制点:从G1进入S和从G2进入M。这两个转变过程都是由一种称为成熟促进因子(maturation-promoting factor,MPF)的蛋白质复合体所触发的(具体调节过程可参阅有关专著)。各个控制点的有序进行能保证通过有丝分裂产生具有正常遗传性能和生理功能的子代细胞。如果调控途径异常,细胞周期不能正常运转,会导致遗传性能紊乱,细胞分裂异常。
在细胞周期进行过程中,发生了极为复杂的生理生化变化,其中最显著的变化是核酸和蛋白质含量的变化,尤其是DNA含量的变化。从图8.4 可以看出,洋葱根尖分生组织中的DNA,在分裂间期的初期,每个细胞核的DNA含量还较少。只有当达到分裂间期的中期,也就是当细胞核体积增到最大体积一半的时候,DNA含量才急剧增加,并维持在最高水平,然后才开始进行有丝分裂。到分裂期的中期以后,因为细胞核分裂为两个子细胞核,所以细胞核的DNA含量大大下降,一直到末期。
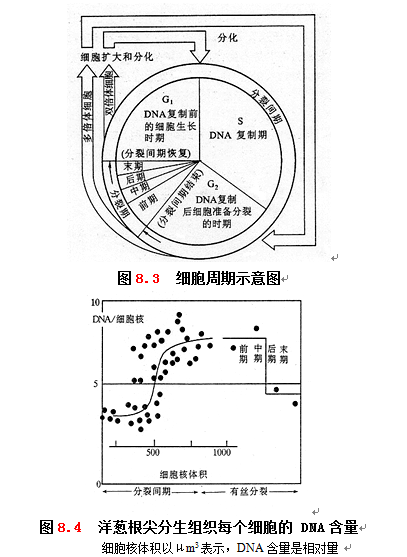
细胞在分裂期的呼吸速率较低,而分裂间期的G1期和G2 期后期呼吸速率都很高。G2 期后期保持较高的呼吸速率,可为分裂期提供充足的能量。
植物激素可影响细胞分裂。实验证明,在小麦胚芽鞘和烟草茎髓的离体培养中,赤霉素可以促进从G1期到S期的过程,从而缩短G1期和S期所需的时间;细胞分裂素促进DNA的合成;生长素是在细胞分裂的较晚时期才起作用,它促进核糖体RNA的形成。
此外,多胺可促进G1期后期DNA的合成,从而促进细胞分裂。维生素,特别是B族维生素,如B1(硫胺素)、B6 (吡哆醇)促进细胞分裂。
不同物种、不同组织的细胞周期及各个分期所经历的时间不同(表8.3)。
表8.3 各种植物分生组织细胞周期及各个分期持续时间(h)
| 植物种类 | G1 S G2 M 全周期 | 温度(℃) |
| 向日葵(Helianthus annuus) 洋葱(Allium cepa) 小麦(Triticum aesticum) 蚕豆(Vicia faba) 玉米(Zea mays) 根尖静止中心 紧邻静止中心的分生组织 | 1.2 4.5 1.5 0.6 7.8 1.5 6.5 2.4 2.3 11.7 0.8 10.0 2.0 1.2 14.0 4.0 9.0 3.5 1.9 24.3
19.3 11.7 3.3 5.1 39.6 2.2 6.1 3.4 2.5 14.2 | 25 24 23 23
23 23 |
8.2.2细胞伸长的生理
根尖和茎端分生组织的细胞分裂形成的新细胞,其体积只有母细胞的一半左右,子细胞必须增大至母细胞同样大小时才能进行下一次分裂。另一方面,在这些子细胞中,除了靠近生长点的一部分仍保留强烈的分生能力外,大多数过渡到细胞伸长时期。
在细胞伸长阶段,细胞的体积显著增加,植物体生长迅速。如豌豆距根尖5~6mm的部位的细胞体积比分生组织的增加20倍。细胞体积的增加,主要是由于水分的进入所造成。由分生组织刚形成的新细胞,细胞体积较小,细胞核相对较大,细胞质浓厚,液泡不明显。细胞进入伸长生长阶段后,细胞中出现小液泡,然后小液泡逐渐增大并合并为一个大液泡,而把细胞核和细胞质挤到边缘。细胞形成液泡后,可进行渗透性吸水,随着水分的进入,细胞体积显著增大。在此过程中,细胞质、细胞核和细胞壁等的干物质都有增加。当细胞体积伸长时,细胞代谢活动十分旺盛,如呼吸速率增快2~6倍,使细胞生长所需的能量供应能得到保证;与此同时,蛋白质合成也显著增加,最高可增大6倍左右(与分生组织细胞相比);二肽酶、蔗糖酶和磷酸酯酶等酶活性加强。呼吸作用的加强和蛋白质合成的增加是细胞伸长的基础。
植物细胞的生长最主要的是基于原生质的增加。由于原生质体外面有细胞壁的限制,如果没有细胞壁相应地延伸,细胞的生长是不可能的。因此,许多学者从细胞壁的物理化学特性来探讨细胞生长的机理。
植物体的分生组织区(如茎尖、根尖)在生长过程中不断产生新细胞。子细胞开始伸长生长时,即为初生壁的形成时期。这些新细胞的细胞壁为初生细胞壁,它比较薄,且具有一定的弹性,以便适应细胞的生长。高等植物的初生细胞壁的基本构成为:以纤维素微纤丝为骨架,以其他两种多糖(半纤维素和果糖)以及糖蛋白为基质,通过共价键和非共价键的结合,交叉形成一个高度复杂的、抗张力强的网状结构。植物细胞壁的基本结构物质主要是纤维素,许多纤维素分子构成微纤丝,细胞壁就是以纤维素微纤丝为基本网状骨架,网眼中充满水、半纤维素、果糖和糖蛋白等。
近年来,对初生细胞壁中的糖蛋白研究较多,特别是伸展蛋白。初生壁中包含多种糖蛋白,其中很重要的一类为伸展蛋白(extensins)。它是一种富含羟脯氨酸的糖蛋白,其分子结构包含两个组成部分:蛋白质骨架和寡糖侧链。蛋白质的氨基酸序列已经被检测,由305个氨基酸组成,分子量约为34kDa,其中含25个Ser-(Pro)4重复序列,其他序列如Try-Lys-Tyr-Lys,Thr-Pro-Val也有多次重复,其中羟脯氨酸残基占30%。伸展蛋白中碳水化合物的主要成分是阿拉伯糖和半乳糖。阿拉伯糖是以寡聚形式与蛋白质骨架上的羟脯氨酸相连结;半乳糖则以O-糖苷键的形式与蛋白质骨架上的丝氨酸相结合。伸展蛋白对细胞壁的弹性张力和韧性起着加强作用。实验证明,当初生壁向次生壁转变时,随着伸展蛋白含量的增加,细胞壁的刚性也增加。在烟草原生质体的培养中,若抑制伸展蛋白的合成,则不能再生正常的细胞壁。这说明伸展蛋白对于初生壁的完整性,以及使其他细胞壁多聚物的正确装配是需要的。此外,近年来研究结果表明,伸展蛋白对植物的抗病性可能具有重要作用。
细胞伸长生长时,即为初生壁的形成时期。当细胞生长接近停止时,才开始产生次生壁。有迹象表明,伸展蛋白在壁中的大量沉积可能是细胞壁木质化的先兆。次生壁除含纤维素和半纤维素外,还常含木质素和栓质等。微纤丝的沉积方向受到质膜下微管排列方向的控制。微纤丝沿细胞纵轴排列,并与纵轴成一定角度,而且,在次生壁的外、中、内三层中,微纤丝的走向也不一致,这样的排列方式大大增强了细胞壁的坚固性。
赤霉素和生长素通过影响细胞壁的可塑性,使细胞壁变得松弛而促进细胞的伸长;脱落酸抑制细胞伸长;细胞分裂素和乙烯则促进细胞扩大。
8.2.3 细胞分化的生理
高等植物个体是不同器官组成的,器官由不同组织构成,而组织又由结构和功能相同细胞构成的。不同器官和组织的细胞具有不同的结构和功能。在个体发育过程中,细胞后代在形态、结构和功能上发生差异的过程称为细胞分化(cell differentiation)。通过细胞分裂和分化,同样来源于分生组织的细胞发育为形态、结构和功能各异的细胞类型。如植物分生组织可分化为不同的组织:薄壁组织、输导组织、机械组织、保护组织和分泌组织等。因此,个体发育是通过细胞分化过程实现的。细胞分裂和细胞分化有着严格的程序和规律。细胞分化过程的实质是基因按一定程序选择性的活化或阻遏,也就是说,细胞分化是基因有选择地表达的结果。
8.2.3.1细胞的全能性
植物细胞全能性(totipotency)的概念是1902年由德国著名植物学家Haberlandt首先提出的。他认为,高等植物的器官和组织可以不断分割直至单个细胞,每个细胞都具有进一步分裂和发育的能力,并提出一个大胆的设想:从一个体细胞可以得到人工培养的胚。具体地讲,植物细胞全能性是指植物体的每一个活细胞都有该物种一套完整的基因组,并具有发育成完整植株的潜在能力。植物体的所有细胞都来源于一个受精卵的分裂。受精卵在不断分裂分化过程中,会形成根、茎、叶等不同器官或组织,但它们都具有相同的基因组成,都携带着亲本的全套遗传特性。因此,只要培养条件适宜,离体培养的细胞就有发育成完整植株的潜在能力。
为了验证自己的设想,Haberlandt也曾作了大量的研究,但由于当时技术和条件的限制,试验均未能成功。随着细胞和组织培养技术的不断发展,在细胞悬浮培养的实验基础上,1958年Steward等利用胡萝卜根的韧皮部组织培养出了完整的新植株。后来,Visil和Hildebrandt又用烟草组织培养的单个细胞培养出了可育的完整植株。1969年Nitch将烟草的单个单倍体孢子培养成了完整的新植株。1970年Steward用悬浮培养的胡萝卜单个细胞也培养成了可育的植株。这些实验结果有力地证明:①高度分化的植物细胞,遗传物质并没有丢失,细胞具有全能性;②在二倍体的同源染色体中只要有一个基因组,即含有该物种的全部基因。
植物细胞全能性的揭示,不仅推动了植物细胞生物学的理论研究,而且为生产实践开辟了广阔的途径。植物组织培养、细胞培养、原生质体培养技术已得到了很大的发展,产生了巨大的经济效益。
8.3.2.2影响分化的因素
细胞分化既受遗传基因的控制又受环境因素的影响。目前,对控制细胞分化的详细机理仍然不清楚,但人们从细胞的体外培养实验中,已经获得了一些线索。
首先,植物激素对细胞分化有重要作用。在植物组织培养过程中,由愈伤组织分化为
根和芽,是由细胞分裂素与生长素含量的比值决定的。CTK/IAA比值低时,促进根的形成;CTK/IAA比值高时,促进芽的形成;两种激素含量相当时,则愈伤组织不分化,继续形成新的愈伤组织。此外,植物激素在维管组织分化中起重要作用。如生长素可诱导愈伤组织分化形成木质部。在丁香愈伤组织中插入一小块丁香的茎尖,发现在接触点之下的愈伤组织里有零散的木质部管胞的分化。如果用IAA代替茎尖,也有相同的现象发生。细胞分裂素在诱导木质部分化过程中与生长素有相似效应。
其次,糖(蔗糖或葡萄糖)浓度与木质部和韧皮部的分化也有关系。在丁香茎髓的愈伤组织培养时,若培养基中糖浓度较低,将诱导形成木质部;若糖浓度较高,将形成韧皮部;若糖浓度在中等水平(2.5%~3.5%),则诱导木质部和韧皮部的同时形成,而且中间还有形成层。
此外,细胞分化受环境因素的影响。光照、温度、营养、矿质元素以及地球的引力等都能影响细胞的分化和促进组织的分化。如光对植物细胞分化的影响就非常复杂:光能影响植物的形态建成,促进组织的分化;光照的长短还能影响植物的开花期;低温处理,能使小麦通过春化而进入幼穗分化;对作物多施氮肥,能使植物营养生长旺盛,延迟开花;由于地球的引力而使植物具有向重力性的特征。
植物体无论是器官或组织水平,还是细胞水平,在形态、结构和生理生化上常常表现出两极差异,这种两极分化的现象就是极性(polarity)(详见本章第5节)。极性是分化的控制因素,还是分化的一种早期表现,仍然是一个争论的问题。
8.3 植物组织培养
8.3.1植物组织培养的概念及类型
植物组织培养(plant tissue culture)是指在无菌和人工控制的环境条件下培养植物的离体器官、组织或细胞的技术。用于离体培养进行无性繁殖的各种植物材料称为外植体(explant)。根据外植体的不同,把组织培养分为五种类型,即愈伤组织培养、悬浮细胞培养、器官培养(包括胚、花药、子房、根和茎的培养等)、茎尖分生组织培养和原生质体培养, 其中愈伤组织培养是一种最常见的培养形式。所谓愈伤组织(callus),原是指植物在受伤之后在伤口表面形成的一团薄壁细胞,在组织培养中,则指在人工培养基上由外植体长出来的一团无序生长的薄壁细胞。除茎尖分生组织培养和一部分器官培养以外,其他几种培养形式最终也都要经历愈伤组织才能产生再生植株。此外,愈伤组织还常常是悬浮培养的细胞和原生质体的来源。
外植体在培养基中培养时,首先脱分化形成愈伤组织,然后再根据不同的实验目的进行愈伤组织培养。在进行愈伤组织培养时,可通过营养物质和生长调节物质的适当配比来诱导愈伤组织分化出芽和根的顶端分生组织,由此产生新的植株。愈伤组织培养物也可放在摇床上,通过试管液体悬浮培养分散成单个细胞。这种单个细胞具有丰富的细胞质、小型的液泡和大的细胞核,具有胚性细胞的特征。这种细胞在适宜的培养基中可发育成胚状体,然后再继续发育成植株。还可通过细胞壁的水解酶(纤维素酶和果胶酶)除去细胞壁,获得原生质体,然后通过原生质体培养也可产生再生植株。广泛的研究结果表明,几乎植物体上所有各部分的组织细胞,包括根、茎、叶、花药、子房及胚珠等,均可通过诱导出来的培养细胞再生新植株。这些结果说明,植物体的每个生活细胞都具有遗传上的全能性。
组织培养的优点:(1)用料少,节约母株资源;(2)繁殖系数高,一块组织或小植株一年内可以繁殖成千上万株小苗;(3)占地面积少,繁殖速度快,在20平方米的培养室中一年可以繁殖30万株试管苗;(4)不受自然气候变化影响,在人工条件下能每年进行大规模生产;(5)生产周期短,繁殖一代小苗仅需一个月左右;(6)可以脱去自然无性繁殖的植物体内所感染的病毒,使之复壮;(7)可意外地在培养中获得突变体、多倍体与有经济价值的新类型;(8)由于组织培养是在试管中进行,携带方便,利于地区间、国际间的种质交流。(9)在引种时许多植物由于地理及气候等原因不能开花结实,可以用组织培养进行营养繁殖。
8.3.2植物组织培养的原理
组织培养的理论依据是植物细胞具有全能性。完整植株中的每个生活细胞保持着潜在的全能性,细胞分化完成后,就受到所在环境的束缚而稳定下来,但这种稳定是相对的。一旦脱离原来所在的环境,成为离体状态时,在适宜的营养和外界条件下,就会表现出全能性,生长发育成完整的植株。因此,在组织培养中就有一个细胞脱分化和再分化过程。所谓脱分化(dedifferentiation),是指已经分化的植物器官、组织或细胞在离体培养时,失去原有的形态和机能,又恢复细胞分裂的能力并形成与原有状态不同细胞的过程。新形成的细胞就是愈伤组织。再分化(redifferentiation)是指脱分化形成的愈伤组织细胞在适宜的培养条件下又分化为胚状体 (embryoid),或直接分化出根和芽等器官形成完整的植株。胚状体具有根、茎两个极性结构,因此可以一次性形成完整的植株(图8.5)。
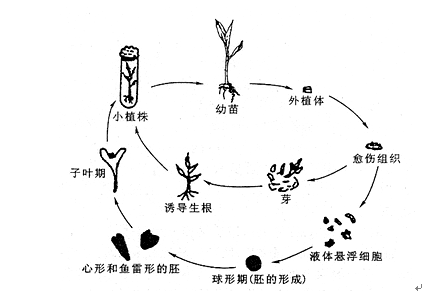
图8.5 植物组织培养的过程
8.3.3植物组织培养的方法
8.3.3.1外植体的选择
植物细胞具有全能性。但全能性表达与否以及表达的难易程度因外植体的生理状态而异。一般来说,与成熟的及高度分化的细胞相比,由分生细胞和胚细胞产生的愈伤组织具有较高的再生能力。受精卵、发育中的分生组织细胞和雌雄配子体及单倍体细胞是较易表达全能性的。由根、下胚轴及茎形成的愈伤组织分化成根的频率很高;由叶或子叶形成的愈伤组织分化成叶的频率很高;由茎端形成的愈伤组织分化成芽和叶的频率亦很高;靠近上部的茎段与接近基部的茎段相比能形成较多的花枝和较少的营养枝。因此,在植物组织培养中,要根据研究目标,有针对性地选择外植体。
8.3.3.2培养基的配制
培养基(medium)中含有外植体生长所需的各种营养物质。不同的外植体、不同的培养方法、不同的培养目的等,都要求采用不同的培养基。现有培养基种类很多,其中White培养基是最早的植物组织培养基之一,其中包含了所有必需的营养成分,被广泛用于离体根的培养;MS ( Murashige和Shoog ,1962 )培养基中含有较高的硝态氮和铵态氮,适合于多种培养物的生长;N6培养基含有与MS差不多的硝态氮,但铵态氮仅为MS的1/4多一些,特别适合于禾本科花粉的培养;B5培养基则适合于十字花科植物的培养;各种培养基适合哪种组织培养可参阅有关文献。通常把大量元素、微量元素、螯合铁和除蔗糖之外有机附加物配成母液,再配制成所需培养基。表8.4 列出目前较常用的几种培养基。
表8.4几种常用培养基的配方(mg/L)
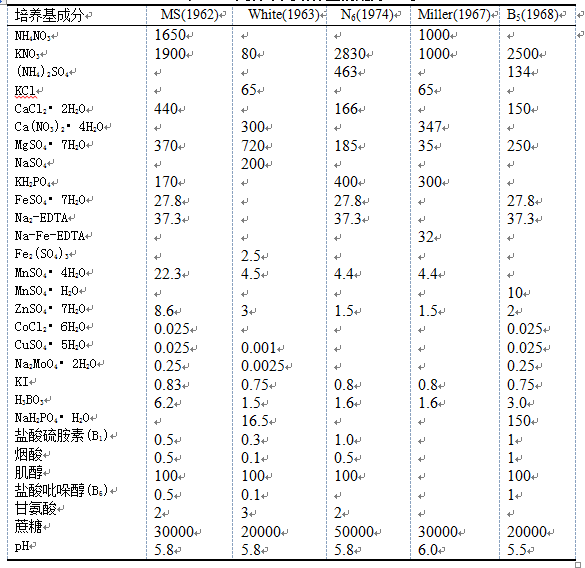
如上所述,表中各种培养基配方虽有所不同,但其主要成分是相同的,都是由五大类物质组成。
1、 机营养物 大量元素和微量元素,即植物生长所必需的N、P、K、Ca、Mg、S、Zn、Fe、B、Cu、Mo、Mn、Cl等。N一般用铵态氮和硝态氮。 当作为唯一的氮源时,硝酸盐的作用要比铵盐好得多,但在单独使用硝酸盐时,培养基的pH值会向碱性方向漂移。若与硝酸盐一起加入少量铵盐,则会阻止这种漂移。因此,有多种培养基中都既含有硝酸盐,也含有铵盐。P常用磷酸盐。S常用硫酸盐。由于铁盐易于形成沉淀,故一般用乙二胺四乙酸二钠(EDTA)与铁盐配制成铁盐螯合物。
2、碳源 一般用蔗糖,其浓度为2%~4%(质量浓度为30~50g/L),有时也用葡萄糖作为碳源。碳源还具有维持培养基渗透压的作用。
3、维生素 硫胺素(B1)是必需的,烟酸、盐酸吡哆醇(B6)、肌醇对生长起促进作用。
4、有机附加物 如氨基酸(甘氨酸)、水解酪蛋白、酵母汁、椰子乳等,它们能促进外植体细胞的分化。但若基本培养基的配方适当,则这些有机附加物在大多数情况下是不需要的。
5、生长调节物质 常用的有生长素类和细胞分裂素类,有时也加入赤霉素。生长素类被用于诱导细胞的分裂和根的分化。常用生长素有IAA(吲哚乙酸)、IBA(吲哚丁酸)、NAA(萘乙酸)、2,4-D(二氯苯氧乙酸)和2,4,5-T (三氯苯氧乙酸),其中IBA 和NAA广泛用于生根,并能与细胞分裂素一起促进茎的增殖。2,4-D和2,4,5-T对愈伤组织的诱导和生长效果明显。生长素一般溶于95%酒精或0.1mol/L的NaOH中。细胞分裂素可以促进细胞分裂和由愈伤组织或器官上分化不定芽。常用细胞分裂素有KT、6-BA、2-iPA(异戊烯基腺嘌呤)、ZT。细胞分裂素一般溶于0.1mol/LHCl或稀薄的NaOH中。与生长素和细胞分裂素相比,赤霉素不常使用。据报道,赤霉素能刺激组织培养中形成的不定胚正常发育成小植株。常用的赤霉素是GA3,赤霉素易溶于水,但溶于水后不稳定,容易分解,实验中常用95%的酒精配成母液在冰箱中保存。
8.3.3.3 灭菌
组织培养是在严格无菌的条件下进行,所以灭菌的好坏就成为成功的关键。常见的消毒灭菌方法有以下几种:
1、化学灭菌法:该法适用于外植体和接种者手等。外界采集的外植体和手的表面携带着各种污染微生物,在把外植体接种到培养基上之前必须进行彻底的表面消毒。常用的消毒剂有:70%酒精、次氯酸钙(漂白粉)、氯化汞(HgCl2升汞)等。消毒所需时间依外植体的种类不同而异,培养材料消毒处理的目的是获得无菌并仍保持着生命力的外植体,因此,应当正确选择消毒剂的浓度和处理时间,以尽量减少组织的伤害。手只能用新洁尔灭等对人体无害消毒剂。
2、高压蒸汽灭菌法:将需要灭菌的物品放在密封的高压高温灭菌锅内,在121℃高温和1个大气压/cm2的压力下,持续15~20分钟,就可杀灭一切微生物的营养体及其孢子。该法适用于培养基、器皿和工具等的消毒灭菌。
3、干热灭菌法:将物品放入烘箱,加热到160℃,持续1~2小时,可使微生物细胞内的蛋白质凝固而达到灭菌的目的。
4、 过滤除菌法:有些试剂或药品受热后不稳定,易分解而失去药效,这类物品的消毒灭菌常采用过滤除菌。将带菌的液体或气体通过孔径小于0.45μm的微孔滤器装置,使杂菌受阻隔留在滤板上,而液体或气体进入无菌瓶内,从而达到除菌目的。
5、 辐射灭菌法:通常用紫外线灭菌。该法适用于接种室等灭毒。
8.3.3.4 接种和培养
植物组织培养是一种无菌培养技术,因此要求工作人员在操作过程中遵守无菌操作规程,一切用具、材料、培养基要求在无菌条件下进行操作。接种时,在接种室的超净工作台上,用无菌镊子将灭过菌的植物组织(材料)从容器中取出,放在无菌培养皿或铺垫上,用无菌解剖刀或剪刀分割植物器官。根据需要切成适当大小(一般外植体为0.5㎝左右)的组织块,再用无菌镊转移接种到预先准备好的培养基中,密封培养瓶(罐)待培养。
无菌培养是将接种在无菌培养基上的外植体,置于培养室的专用培养架上培养,要求培养室里清洁少菌,调控温度、光照和通气等环境因子。植物组织培养通常采用的培养温度为23~28℃;采用的照光强度在3~90 mmol·m-2·s-1,一般用9 mmol·m-2·s-即可,照光时间和强度根据实验目的而定,有连续照光或周期性照光均可,光质通常采用的是荧光灯;组织培养在固体培养基上,培养瓶内的空气成分及通气量就能满足培养物的生长,但用液体培养基时,特别是细胞悬浮培养,一般用震荡法通气,震荡速度约为100~150 rpm,也可根据不同培养材料所要求而定。
8.3.4组织培养的应用
植物组织培养的应用十分广泛,主要表现在以下领域:
1.无性系的快速繁殖。
用组织培养法快速繁殖园艺作物,是组织培养应用最有成效的实例,首先在兰花上成功应用。自Morel在1960年得到兰花组织培养苗后,很快应用于生产,形成了组织培养繁殖兰花工业。目前可用组织培养繁殖的兰花已有数百种,同时在主要经济作物甘蔗、菠萝、香蕉、草莓、球茎花卉等多种作物繁殖种苗上,已成功应用。
2.培养无病毒种苗。
植物病毒是目前无法治疗的病毒,一般只能采用防治蚜虫传播的间接措施,农作物中很多植物都带有病毒,特别是无性繁殖植物,像马铃薯、草莓、大蒜、康乃馨等等,常常是带着病毒代代相传,造成产量降低、品质劣化。用茎尖培养可以得到无病毒植株,用它作为继续繁殖的母株,就可以得到大量无病毒幼苗。实践证明,用这种方法可以除去马铃薯携带的Y、A、M、S病毒和奥古巴花叶病毒。脱毒植株的产量明显比感病植株高。脱毒大蒜的蒜头比感病的大得多。此法已在草莓、葡萄、康乃馨、百合等多种作物上获得成功,产生了明显的经济效益。
3.新品种的选育:
⑴.花培和单倍体育种。
自1964年Guba等培养曼陀罗花药获得了单倍体植株以来,各国科学家都致力于花药培养,1980年就有23个科、52个属、160多个种的植物,通过花药培养获得单倍体植株,到目前已有数百种植物花药培养成功。我国科学家在单倍体育种方面作出了杰出贡献。1974年用单倍体育种法育成世界上第一个作物新品种——单育1号烟草品种。随后又育成大面积栽培的作物新品种,如水稻中花8号、小麦京花1号等等。
⑵.离体胚培养和杂种植株获得。
在远缘杂交中,杂交后形成的胚珠往往在未成熟状态时,就停止生长,不能形成有生活的种子,因而杂交不孕,给远缘杂交造成极大困难。在十九世纪二十年代末,Laibanch用胚培养技术培养亚麻种间杂交胚,第一个获得了杂种植物,这为远缘杂交时克服杂交不亲和的障碍提供了一项有用的技术。这项技术发展至今,已相当成熟,可以说多数植物的成熟或未成熟胚通过培养都可获得成功。幼胚培养可使约50个细胞大小的极幼龄的胚状结构培养成植株。远缘杂交中,由于生理上和遗传上的障碍而不能杂交成功,可采用试管受精加以克服,即将母本胚珠离体培养,使异种花粉在胚珠上萌发受精,产生的杂种胚在试管中发育成完整植株。用此法已获得甘蓝属种间杂种。
⑶.体细胞诱变和突变体筛选。
培养细胞处在不断分生状态,它容易受培养条件和外加压力(如射线、化学物质)的影响而产生诱变,从中可以筛选出对人们有用的突变体,从而育成品种,目前用这种方法筛选到抗病、抗盐、高赖氨酸、高蛋白、抗除草剂的突变体,有的已用于生产。
⑷.细胞融合和杂种植株的获得。
通过原生质体融合,可部分克服有性杂交不亲和性,而获得体细胞杂种,从而创造新种或育成优良品种。自1972年获得烟草第一个种间体细胞杂种以来,即在多种植物上进行试验,已获得种内、种间和属间杂种植物,但这些杂种尚未能表现实际应用价值,有待进一步研究解决稳定性等问题。
4.人工种子和种质保存。
1978年,Murashige首次提出了人工种子的概念,即利用植物组织培养具有体细胞胚胎发生的特点,把胚状体包裹在胶囊内形成球状结构,使其具有种子的机能并可直接播种田间。人工种子(artificial seed)又称人造种子,它包括人工胚状体类培养物、人工胚乳和人工种皮三个部分。Kitto和Janick用聚氧乙烯包埋胡萝卜胚状体,首次制成了人工种子。人工种子具有巨大的应用潜力,它在快速繁殖优良品种与无性系,固定杂种优势、简优育种程序及基因工程,在去病毒技术及其它生物技术相结合等方面,均有非常诱人的前景。当然,目前该技术尚处于发展研究阶段。
种质资源的保存引起科学家和各国政府的极大重视,因为世界种质资源日益枯竭,大量有用基因损失,特别是那些不产籽或种子寿命短的生物更为严重。利用组织和细胞培养法低温保存种质,给保存和抢救有用基因带来希望和可能,体外培养物在-20℃至-196℃的低温下贮藏数月甚至更久,尚能恢复生长,再生成植株。
5.药用植物和次生物质的工业化生产。
植物几乎能生产人类所需要的一切天然有机化合物,如蛋白质、脂肪、糖类、药物、香料、生物碱及其他天然活性物质,而这些化合物都是在细胞内合成的。利用次生物质的细胞工程来开发天然植物资源,可以克服植物本身有用成分含量低,生长速度慢,种群稀少等造成的资源紧缺现状,国际上植物次生物质细胞工程研究已有四十年的历史,迄今为止从培养细胞中分离的次生物质近600多种,其中有近40种成分超过了原植物体含量,紫草细胞有效成分紫草素含量达细胞干重的10%以上,像人参、紫草、黄连、毛地黄等细胞培养生产其有用药用成分已实现了工业化生产。
8.4 植物的生长
8.4.1植物生长的周期性
在自然条件下,植物或植物器官的生长速率随昼夜或季节发生有规律的变化,这种现象叫做植物生长的周期性(growth periodicity)。植物生长的周期性包括昼夜周期性(温周期性)和季节周期性。
8.4.1.1 植物的生长曲线和生长大周期
根、茎、叶、种子、果实等器官以及一年生植物的整株植物,在整个生长过程中,生长速率都表现出“慢—快—慢”的特点,即开始时生长缓慢,以后逐渐加快,达到最高速度后又减慢以至最后停止。以植物(或器官)体积对时间作图,可得到植物的生长曲线。生长曲线表示植物在生长周期中的生长变化趋势,典型的生长曲线呈“S”形(图8.6)。植物体或个别器官所经历的“慢—快—慢”的整个生长过程,称为生长大周期(grand period of growth)。
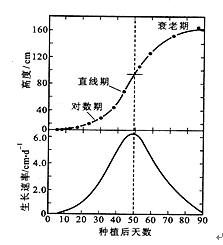
图8.6 玉米的生长曲线
植物个体生长是以器官生长为基础,器官生长是以细胞生长为基础,由于细胞生长速率存在“慢—快—慢”的变化规律。即细胞处于分裂时期,虽然细胞数量多但体积小,因此生长缓慢;细胞进入伸长期,由于液泡的出现并不断合并使体积迅速增大,所以生长迅速;而细胞处于分化期,因此体积基本定型,故生长极为缓慢乃至停止,从而使个体与器官的生长同样表现出这种规律。此外,一年生植物的生长速率表现出生长大周期的原因是由于生长初期植株幼小,合成干物质的量少,因而生长缓慢;以后因为枝繁叶茂,光合作用大大加强,合成大量有机物,干重急剧增加,因而生长加快;最后由于植株进入衰老期,光合作用下降,合成有机物的量减少,加上呼吸作用的消耗,因此,植株生长缓慢以至停止。
掌握植物一生生长速率的变化规律,有利于促进或控制植物的生长。农作物的前期生长缓慢,有延迟期。在农业生产上,可以采取以肥水及其他农业措施来促进营养体的生长,达到早发快长的目的。而作物生长的中期是生长直线上升或指数增长期,任何促进或控制植物生长的措施都必须在生长速率达到最高前施用,才能有效。如要促进作物稳定生长,需要及时供应肥水;如要防止作物过度生长,则需要控肥水供应或使用矮壮素等措施。后期作物生长缓慢,要适当供应肥料,以防止早衰,但又不能过量施肥,以防止植株贪青晚熟。
8.4.1.2生长的温周期性
影响植物生长的因子主要是光照强度、温度和植物体内的水分状况。在地球自转的二十四小时中,昼夜光照强度不同,温度也不同,因而植物的生长速率也不同,表现出生长的昼夜周期性变化。植物的生长随温度的昼夜周期性而发生有规律的变化的现象,被称为植物生长的温周期性(thermoperiodicity of growth),或植物生长的昼夜周期性(daily periodicity of growth)。
植物生长产生昼夜周期性的原因主要是由于昼夜的温度、水分和光照的不同。一般来说,在夏季,植物生长速率白天较慢,夜晚较快。因为白天温度高,光照强,蒸腾量大,植物易缺水,此外,强光会抑制植物细胞的伸长;晚上温度降低,呼吸作用减弱,物质消耗减少,积累增加;较低的夜间温度还有利于根系的生长以及细胞分裂素的合成,从而有利于植物的生长。但在冬季,植物在白天的生长速率比夜晚快,这是由于冬季的夜晚温度太低,植物的生长受阻。
8.4.1.3生长的季节周期性
植物在一年中的生长速率,随季节变化而发生有规律性的变化,称为植物生长的季节周期性(seasonal periodicity of growth)。这是因为一年四季中,光照强度、温度和水分等影响植物生长的外界因素是不同的。在温带地区,四季分明。春天,气温回升,日照延长,植株上的休眠芽开始萌动,长出茎、叶;夏天时,温度与日照进一步升高和延长,水分较为充足,植株进行旺盛生长;秋天时,气温逐渐下降,日照逐渐缩短,植株的生长速率逐渐减慢、停止,叶子脱落,进入休眠状态;到了冬天,植株处在休眠状态下。次年春季又开始生长,周而复始。
生长的季节性变化是建立在体内代谢活动的基础之上的。当秋天来临时,日照长度缩短,这个信号被叶片感受后,经信号转导产生一系列代谢变化,导致植物对冬季的气候产生种种生理上的适应,如物质从叶片转移到根、茎和芽中储藏起来,体内糖分与脂肪等物质的含量提高,组织含水量下降,原生质转为凝胶状态,植物抗性增强;生长素、细胞分裂素、赤霉素由游离态转变为束缚态,脱落酸等抑制生长的激素逐渐增加,体内代谢活动大大降低,生长停止,进入休眠状态。进入第二年春季后,内源激素发生变化,休眠逐渐解除,恢复生长。
植物的生长习性使植物体内营养物质生产、分配、再分配和再利用,有着一个动态的变化。多年生木本植物尤为明显。春、夏季节,植物主要依靠当时光合作用生产的有机物供应茎、叶、花和果实的生长,秋季将营养物质储藏到根、茎和芽中,次年又利用它们供生长之用。所以,从植物体内物质分配、利用和储藏以及不同器官的生长状况,也可以看出植物生长的季节周期性变化(图8.7)。
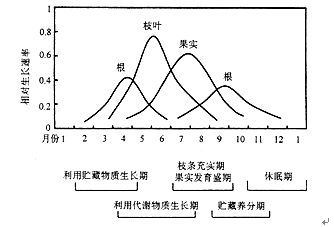
图8.7 梨树周期性生长动态示意图
此外,植物的年轮也是植物生长季节周期性的一个具体表现。年轮是由于形成层在不同季节所形成的次生木质部在形态上的差异而形成的。在同一圈年轮中,由于春夏季的气温适于树木生长,形成层的活动旺盛,所形成的木质部细胞较大,细胞壁较薄,因而木材质地疏松,被称为“早材”;到了秋冬季,由于气候逐渐干冷,形成层活动逐渐减弱以至停止,所形成的木质部细胞小,细胞壁厚,木材质地紧密,颜色较深,被称为“晚材”。前一年的晚材和第二年的早材界限分明,此即年轮线。
8.4.2影响植物生长的外界条件
植物的生长是在各种生理活动协调进行的的基础上细胞分裂和生长的结果。这些生理活动包括光合、呼吸、水分的吸收与蒸腾、矿物质的吸收与转运、有机物的转化与运输等过程。因此,凡能影响这些生理活动的外界条件都能影响植物的生长,主要包括温度、光照和水分。此外,矿质元素和植物生长调节剂对植物生长起调节作用。
8.4.2.1温度
温度对植物生长的影响是通过酶而影响各种代谢过程的一种综合效应。温度不仅影响水分与矿质的吸收,而且影响物质的合成、转化、运输与分配,因而影响细胞的分裂与伸长。在一定温度范围内,温度升高植物生长加快,温度降低植物生长减慢。每种植物的生长都有温度三基点,即生长的最低温度、最适温度和最高温度。温度三基点与植物的原产地有关(表8.5 ),原产热带及亚热带的植物,其温度三基点较高,而原产温带的植物,其温度三基点较低。
表8.5几种农作物生长的温度三基点(引自曾广文等,2000)
| 作物 | 最低温度/℃ | 最适温度/℃ | 最高温度/℃ |
| 水稻 大小麦 向日葵 玉米 大豆 南瓜 棉花 | 10~12 0~5 5~10 5~10 10~12 10~15 15~18 | 20~30 25~31 31~37 27~33 27~33 37~44 25~30 | 40~44 31~37 37~44 33~50 33~40 44~50 30~38 |
生长的最适温度是指植物生长最快的温度,但并不是植物生长最健壮的温度,因为在最适温度下,由于植物体内的物质消耗太多,因此常比最适温较低温度下的生长弱,这种对植物生长最健壮而且比最适温度稍低的温度称为协调最适温度。植物生长的温度范围是比较窄的。在生长的最低温度与维持生命的最低温度之间,以及生长最高温度和维持生命的最高温度之间,新陈代谢活动仍能进行,但生长完全停止了,这是因为生长是一个复杂的过程,是体内各种生理活动协调统一的结果,超过一定温度范围后,代谢活动虽然能够进行,但整体的协调已被破坏。
植物不同生育期对温度的要求不同。一年生植物从种子萌发到开花结实各个时期所要求的温度,一般正好和自然界从春季到秋季的气温变化相吻合。因此,作物栽培要注意适时播种。从外地引种时,也必须注意温度条件是否适合。
植物不同器官生长的温度范围也有区别。温带木本植物根生长的最低温度在2~5℃,较枝条生长所要求的温度低。春季枝条从芽中抽出以前,根已开始生长,晚秋地上部分停止生长后,根仍可继续生长。
如前所述,植物的生长还具有温周期性。我国劳动人民很早就已注意到夜间降温的有利作用,这就是温周期现象在生产实践中的应用。
在农业、林业和园艺生产中,了解各种植物的生长对温度条件的要求,在温室栽培中,调节昼夜温度的变化,对于使栽种作物能正常生长发育、提高产量,具有重要的意义。
8.4.2.2 光照
光对植物生长的影响可分为间接作用和直接作用两个方面。
光对植物生长的间接作用,是指光通过影响光合作用、蒸腾作用和物质运输等,从而影响植物的生长。这个间接作用是一种高能反应,因为光是光合作用的能源,光照不足就不能产生足够的有机物,植物生长也就失去物质基础。此外,光还可以影响植株的蒸腾作用,在土壤水分不足的情况下,往往会使植物体内产生水分亏缺而抑制植物的生长。光对植物生长的直接作用是指光形态建成作用(详见7.植物的光形态建成)。黑暗中生长的幼苗与光下生长的幼苗在形态上有很大的差异。黑暗中生长的幼苗,植株瘦长,茎细长而脆弱、机械组织不发达、顶端呈弯钩状、节间很长,叶片细小、不能展开、无叶绿素、不能进行光合作用,同时根系发育不良。由于黑暗中生长的幼苗茎、叶均为黄色,因而被称为黄化苗(图8.8)。
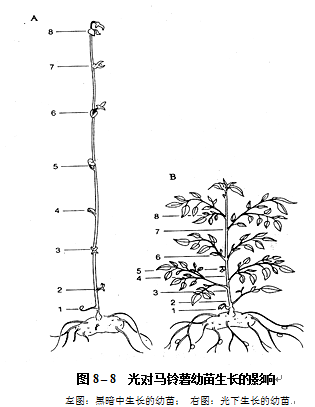
黑暗中生长产生黄化苗的现象被称为黄化现象(etiolation)。红光促进幼叶的展开,而抑制茎的过度伸长,对消除黄化现象起着最有效的作用。
了解光对植物生长的影响,对于农业生产实践具有指导意义。在作物栽培中,要合理密植。因为栽种过密会导致群体内光照减弱,一方面影响光合作用,另一方面还使茎秆长得细长,容易倒伏。黑暗中培养韭黄、蒜黄和豆芽的栽培技术,正是利用黄化现象。又如在低温下塑料薄膜覆盖育秧,浅蓝色薄膜的效果比无色的好,秧苗茁壮,插秧后成活快。这是因为,一方面浅蓝色薄膜能吸收大量600nm的橙光,使膜内温度升高,有利于秧苗生长;另一方面浅蓝色薄膜能透过400~500nm的蓝紫光,抑制秧苗生长,使幼苗茁壮。
8.4.2.3水分
水是原生质的重要组成部分,原生质平均含水量80%~90%。水是生物化学反应的必要介质,细胞内各种代谢活动都是在水溶液中进行。根系从土壤中吸收的矿质元素都是以水溶液状态被吸收的。植物与环境间气体交换也是以水溶液状态出入细胞,植物体内的物质运输以液流方式进行。因此,水分在植物的生命活动中具有十分重要的意义,水分状况对植物的生长也有重要的影响。
植物要进行正常的生长活动,细胞原生质必须处于水分饱和状态。不论细胞的分裂、生长和分化,还是组织、器官和个体的生长,均需有充分的水分供应。植物的不同组织、器官的结构和代谢功能不同,以及不同植物对环境的适应性不同,从而对水分的需要量也不同,但充分又适当的水分供应,能促进植物的代谢和生长。掌握植物的不同器官、不同组织等对水分需求的差异,就可通过调节水分以控制生长。例如细胞分裂和伸长都必须在水分充足的情况下才能进行。当水分不足时,两者均受抑制。但对细胞伸长的抑制更大。因为分生组织细胞是生长中心,代谢旺盛,有机物含量丰富,水势低,当植物体水分不足时,分生细胞将向其它细胞吸收水分,以满足自身代谢的需要;而细胞的伸长生长是以液泡吸水为基础的,当缺水时,细胞的伸长生长明显受到抑制。利用这种差异,可在稻麦拔节时控制水分的供应,让稻田排水可控制茎基第一、二节的延长以防止倒伏。同样,在水稻催芽时,常常用控制水分供应来限制种苗的生长,避免种苗过大,播种时易伤苗。相反,水稻、小麦等禾谷类作物从分蘖末期到抽穗期为第一个水分临界期,这时缺水将严重影响到穗轴的伸长,阻碍正常的抽穗而导致减产。
8.4.2.4矿质元素
土壤中含有植物生长必须的矿质元素。这些元素中有些是原生质的基本成分,有些是酶的组成或激活剂,有些能调节质膜的透性,并参与缓冲体系以及维持细胞的渗透势。植物缺乏这些元素便会引起生理失调,影响生长发育,并出现特定的缺素症状。土壤中的矿质元素分为必需元素、有益元素和有害元素,其中前二者促进植物生长,有害元素则抑制植物生长。如氮肥能使出叶期提早、叶片增大和叶片寿命相对延长。但施用过量,叶大而薄,容易干枯,寿命反而缩短。氮肥同样显著促进茎的生长,氮肥过多,会引起徒长倒伏(详见2.植物矿质营养)。
8.4.2.5植物生长调节剂
生长调节物质对植物的生长具有显著的调节作用(详见6.植物生长物质)。GA3能显著促进茎的伸长生长。GA3 对水稻茎秆的节间伸长有非常明显的效果,因此生产上利用赤霉素促进杂交水稻亲本的抽穗,减少包颈率,便于亲本间传粉。生长延缓剂CCC等抑制菊花近顶端分生组织的细胞分裂和茎的生长,外施GA3 可抵消CCC的抑制效果。
8.5植物生长的相关性
高等植物体是由各种器官组成的统一的有机体。构成植物体的各个部分,既有精细的分工又有密切的联系,既相互协调又相互制约,这种植物体各部分间的相互协调与制约的现象称为相关性(correlation)。植物生长的相关性包括地下部与地上部的相关、主茎与侧枝的相关、营养生长与生殖生长的相关等。
8.5.1地下部与地上部的相关
8.5.1.1地下部与地上部的相关表现
地下部是指植物体的地下器官,包括根、块茎、鳞茎等,而地上部是指植物体的地上器官,包括茎、叶、花、果等。地下部与地上部的相关性常用根冠比(root/top ratio, R/T比)表示,即地下部分的重量与地上部分的重量的比值。植物的地上部分和地下部分各处在不同的外部环境中,地上部分所处的环境可以使植物获得充足的阳光、空气,而地下部分可从土壤中吸取足够的水分和矿质元素,两者之间通过维管束进行营养物质与信息物质的交换。根部的活动和生长有赖于地上部分所提供的光合产物、生长素(IAA)和维生素B1等;而地上部分的生长和活动则需要根系提供水分、矿质盐、部分氨基酸以及根中合成的植物激素(CTK、GA、ABA)等,通过物质的交换使两部分的生长相互依存,缺一不可。一般而言,植物根系发达,地上部分才能很好地生长。所谓“壮根必须先壮苗”、“根深叶茂”和“本固枝荣”等民谚深刻地说明植物地上部分和地下部分相互促进协调生长的关系。但在水分、养料供应不足的情况下,常常由于竞争而相互制约。
对于植物来说,根冠比能反映植物的生长状况以及环境条件对植物地上部和地下部生长的不同影响。当环境条件发生变化时,植物地下部和地上部的生长就会发生变化,从而改变了根冠比。对于一定的植物体或一定的生长发育阶段,根冠比保持一定的数值。影响根冠比的因素很多,主要有以下几方面:
1、土壤水分 土壤水分缺乏对植物地上部的影响远比对地下部的影响要大。这是因为,虽然根和地上部的生长都需要水分,但由于根生活在土壤中容易得到水分,而地上部分是植物体散失水的部位而其水分要靠根来供应的,所以缺水时地上部会更早更严重,这时地上部的生长会受到一定限制,根的相对重量增加,而地上部的相对重量减少,使根冠比增大。相反,当土壤水分过多时,由于土壤通气性不良,氧气含量减少,根的生长受到一定程度影响,而地上部由于水分供应充足而保持旺盛生长,因而根冠比减少。水稻生产上出现“旱长根、水长苗”现象,就是这个道理。
2、温度 低温可使根冠比增大,这是因为根部的活动和生长所需要的温度比地上部低一些。在气温低的秋末至早春时期,植物地上部分的生长处于停滞期,而根系仍有生长,因此根冠比增大;当气温升高,地上部分的生长加快,根冠比就下降。
3、光照 在一定的范围内光照加强,光合产物积累增多,地下部的碳水化合物供应得到改善,因而促进根的生长,使根冠比增大。光照不足时,地上部向下输送的光合产物减少,影响根部生长,而对地上部分的生长相对影响较小,因此根冠比降低。
4、矿质元素 不同营养元素或不同的营养水平,对根冠比的影响有所不同。当土壤中氮素缺乏时,地上部比地下部更缺氮,因而地上部的生长受到抑制,根冠比增大;当土壤中氮肥充足时,有利于地上部蛋白质的合成,茎叶生长旺盛,同时消耗较多糖类,使运到地下部的糖类减少,因而根的生长受到抑制,根冠比降低(表8 – 6 )。磷、钾肥有调节碳水化合物转化和运输的作用,可促进光合产物向根和贮藏器官的转移,通常能使根冠比增大。
表8–6 氮素水平对胡萝卜根冠比的影响

5、生长调节剂 整形素、矮壮素、缩节胺、三碘苯甲酸、PP333等生长抑制剂或生长延缓剂对茎的顶端或亚顶端分生组织的细胞分裂和伸长有抑制作用,使节间变短,可增大植物的根冠比。赤霉素、油菜素内酯等生长促进剂,能促进叶菜类如芹菜、菠菜、苋菜等茎叶的生长,使根冠比降低,从而提高作物产量。
在农业生产上,常通过水肥措施来调控作物的根冠比,促进收获器官的生长,以达到增产的目的。对收获器官是地下部分的作物如甘薯、胡萝卜、甜菜、马铃薯等,前期应保证充足的水肥供应,以促进茎叶的生长,增加光合面积,多合成光合产物;而在后期则应减少氮肥和水的供应,增施磷、钾肥,以利于光合产物的向下运输及淀粉的积累,从而促进这些作物的增产。
8.5.1.2地下部与地上部之间的信息转递
植物的地下部与地上部之间除了进行物质和能量的交换外,各部分之间还进行着信息的传递。图8 – 9 概括了土壤干旱时根冠间的物质和信息交流。其中ABA被认为是一种逆境信号,在水分亏缺时,根系快速合成并通过木质部蒸腾流将ABA运输到地上部分,调节地上部分的生理活动,如缩小气孔开度,抑制叶的分化与扩展,以减少蒸腾来增强植物对干旱的适应性。植物地上部也合成一些微量活性物质会影响根系的生理功能。如叶片的水分状况信号,如细胞膨压,以及叶片中合成的化学信号物质(IAA)也可以转递到根部,影响根的生长。除了ABA外,根系合成的细胞分裂素以及氨基酸在这一过程中也可能起着某些作用。除根冠间有化学信号的转递以外,有报道指出根冠间还有电信号转递,目前关于根冠间的信息转递及其相互影响的研究正方兴未艾。
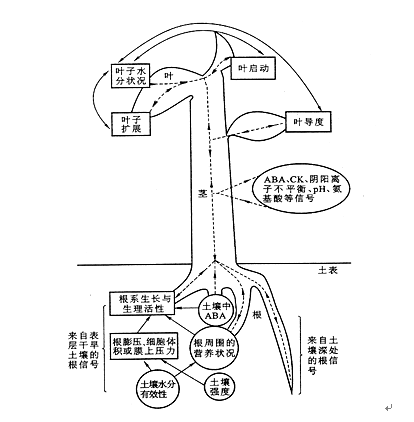
图8–9 土壤干旱时根中化学信号的产生以及根冠间的相关性
(引自W. J. Davies等1991)
←‐→代表化学信号的传递,圆圈代表土壤作用,矩形代表植物生理过程
8.5.2主茎与侧枝的相关
植物的顶芽长出主茎,侧芽长出侧枝,通常主茎生长很快,而侧枝或侧芽则生长较慢或潜伏不长。这种由于植物的顶芽生长占优势而抑制侧芽生长的现象,称为顶端优势(apical dominance或terminal dorminance)。
顶端优势现象普遍存在于植物界。在木本植物(杉树、桧柏)中,由于顶芽长得很快,而侧芽长得慢,树冠呈宝塔形。在草本植物中的双子叶植物(如烟草、油菜、向日葵)中,顶端优势也很显著。不同的植物,顶端优势的强弱有很大差别。例如禾谷类作物中,玉米的顶端优势较强,所以不易分枝;而稻、麦的顶端优势较弱,可以长出多级分蘖。
关于顶端优势产生的原因,很早就引起了学者们的重视,一般都认为这与营养物质的供应和内源激素的调控有关。有多种假说用来解释植物顶端优势。
最早是K. Goebel于1900年提出的“营养假说”,认为顶端构成营养库,垄断了大部分营养物质。顶芽分生组织比侧芽分生组织先形成,具有竞争优势,能优先利用营养物质并优先生长,侧芽因缺乏营养物质而生长受到限制。从形态解剖结构看,侧芽与主茎之间没有维管束的连接,不易得到充分的营养供应,而顶芽由于输导组织发达,成为生长中心,因而竞争营养的能力强。用亚麻试验表明,缺乏营养物质时亚麻的侧芽生长完全被抑制,而营养充足时侧芽可以生长。营养假说未涉及植物激素对生长的调节作用。
1934年K. V. Thimann 和F. Skoog提出的“生长素假说”,认为顶端优势是由于生长素对侧芽的抑制作用而产生的。植物顶端形成生长素,通过极性运输到侧芽,侧芽对生长素敏感性比顶芽强,而使侧芽生长受到抑制。距离顶芽愈近,生长素浓度愈高,对侧芽的抑制作用愈强。其最有力的证据是,植物去顶芽后,可导致侧芽的生长;使用外源的生长素可代替植物顶端的作用,抑制侧芽的生长(图8 – 10 )。另外,施用生长素运输抑制剂,或对主茎作环割处理,阻止生长素的运输,可导致处理部分下方的侧芽生长。
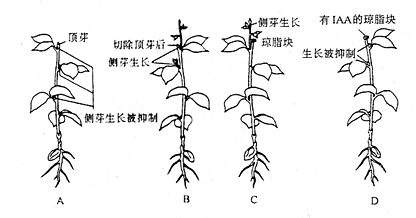
图8–10 生长素与植物的顶端优势
A.有顶芽时,侧芽生长被抑制;B.切除顶芽上部,侧芽萌发生长,并代替顶芽抑制下部侧芽生长;C.切除顶芽,并在切口处放上琼脂,生长情况同B;D.在切口处放上含IAA的琼脂,即使没有顶芽,含IAA的琼脂也抑制侧芽生长
1936年Went提出营养物质定向转移假说来解释顶端优势产生的原因。该学说认为生长素既能调节生长又能影响营养物质的运输方向。植物顶端是生长素的合成部位,高浓度的IAA使顶端成为生长活动中心和物质交换中心,并能将营养物质调运至茎端,因而不利于侧芽的生长。许多实验也证明,植物顶端产生的生长素可以决定矿质元素和同化物在植物体的运输方向及其分布,生长素通过影响同化物在韧皮部的运输来控制植物茎中的营养梯度。
近年来的研究表明,植物顶端优势的存在是某些特定基因控制的多种内源植物激素相互作用的结果。因为这些基因的沉默会导致顶端优势的丧失和产生大量侧芽。实验证明,用细胞分裂素处理侧芽,可以解除顶端对侧芽的抑制作用,使侧芽在有顶端存在的情况下也可萌发生长。因此认为,侧芽生长受抑制是因为根部合成的细胞分裂素优先供应给顶芽,而侧芽缺少细胞分裂素所致。赤霉素有加强植物顶端优势的作用,用赤霉素处理顶芽,可加强对侧芽的抑制作用,但在顶芽被去除的情况下,赤霉素不能代替生长素来抑制侧芽的生长,相反会引起侧芽的强烈生长。
对植物顶端优势的研究,虽然提出了众多假说,但有一点是共同的,即都认为顶端是信号源。这信号源是由顶端产生并向下极性运输的生长素,它直接或间接地调节着其他激素、营养物质的合成、运输与分配,从而促进顶端生长而抑制侧芽生长。而细胞分裂素等其他植物激素、营养物质以及Ca2+浓度等也影响着顶端优势,因此顶端优势可能是多种因子综合影响的结果。
在生产上,常常利用顶端优势的原理来控制植物的生长。我们有时需要利用和保持植物的顶端优势,如麻类、向日葵、玉米、高粱、烟草等作物以及用材树木,需抑制其侧芽生长;有时则需消除顶端优势,以促进分枝生长。如棉花整枝、果树修剪等,就是破坏顶端优势来合理分配养料。棉花在不整枝时,养分运到主茎及营养枝上较多,果枝得不到足够养分,蕾铃就会脱落。整枝后,可以重新分配植株体内的养分,使它集中到果枝上,减少枝叶徒长,促进光合作用正常进行,因而可以避免蕾铃脱落,提高棉花产量和质量。在果树栽培上,正确的修剪枝条非常重要。在花卉栽培上,也可以用打顶或去蕾的方法类来控制花的数目和大小。有时,也可以利用植物生长调节剂代替打顶,如三碘苯甲酸处理大豆,可去除顶端优势,增加分枝,促进多开花结荚。
植物的根也有顶端优势,主根生长很旺,使侧根生长受到抑制。在主根受损时,侧根才较快生长。树苗、菜苗移栽时,主根被截断,可使侧根生长加快,这有利于吸收水分和肥料。实验表明,根的顶端优势可能与细胞分裂素有关,根尖合成细胞分裂素并向上运输,抑制侧芽的生长。脱落酸和黄质醛也能抑制侧根的生长,但要求的浓度较细胞分裂素为高。
8.5.3营养生长与生殖生长的相关
营养生长和生殖生长是植物生长周期中的两个不同阶段,通常以花芽分化作为生殖生长开始的标志,但这两个阶段并不能彼此截然分开。在植物一生中,只一次开花结实植物如水稻、小麦、玉米、高粱、向日葵、竹子等,它们的营养生长和生殖生长阶段一般明显地前后分开。这类植物开花后,营养器官所合成的有机物,主要向生殖器官转移,营养器官逐渐停止生长,整个植株走向衰老死亡。另一类植物,一生能多次开花结实如棉花、番茄、瓜类以及多年生果树等植物,营养生长和生殖生长不能截然分开,而是有所重叠,即在开花阶段,营养生长仍在继续进行。开花结果并不马上引起营养器官的衰老、死亡。多年生果树从种子萌发到开始花芽分化之前的时期为营养生长期,以后便进入营养生长和生殖生长的并进阶段,而且可持续多年。
植物的营养生长和生殖生长之间是既相互依赖又相互制约的关系。营养生长为生殖生长奠定了物质基础。生殖器官生长所需要的养料,大部分是由营养器官所供给的,因此,营养器官生长的好坏直接关系到生殖器官的生长发育。营养器官生长不好,生殖器官的生长发育就不好。另一方面,在生殖生长过程中,生殖器官会产生一些激素类物质,反过来影响营养器官的生长。
营养生长和生殖生长之间还存在着相互制约的关系。营养器官生长过旺,会消耗过多的养分,影响生殖器官的生长发育。如水稻、小麦若前期肥水过多,造成茎、叶徒长,会延迟幼穗分化,显著增加空瘪粒;后期肥水过多,则造成贪青迟熟,影响粒重。又如果树、棉花、大豆等,若枝叶徒长,会造成不能正常开花结实,严重的甚至落花落果。相反,生殖器官的生长也会抑制营养器官的生长。如番茄,在自然状态下开花结实后,让果实自然成熟,营养器官的生长就日渐减弱,最后衰老死亡;如果不断摘除花、果,则营养器官就可以继续旺盛生长。
由于开花结果过多而影响营养生长的现象在农业生产上经常遇到,如苹果、梨、荔枝和龙眼等果树,常发生一年产量高,次年产量低的大小年现象。当果树结实过多时,花、果消耗了大量的养分,致使枝叶中储备的养料不足,花芽分化受阻,来年花果减少。相反,在小年结实少,消耗养分少,枝叶积累营养较多,可形成较多花芽,来年开花结果就多。另外,大小年现象与果树中激素的变化有关,特别是赤霉素含量的变化会影响花芽的分化。因此,在果树生产中适当疏花疏果,可以使果树营养分配方面收支平衡,以便消除大小年,使果树年年丰收。
8.5.4植物的极性与再生
极性(polarity)是指植物体在器官、组织甚至细胞中在不同轴向上存在某种形态结构和生理生化上的梯度差异。主要表现在细胞质浓度的不一,细胞器数量的多少,核位置的偏向等方面。极性的建立会引发不均等分裂,使两个子细胞的大小和内含物不等,由此引起分裂细胞的分化。因此,极性是植物细胞分化的基础。植物体的极性在受精卵中已形成,受精卵第一次分裂所形成的基细胞及顶端细胞即进入不同的发育途径,由基细胞分裂形成高度分化的胚柄细胞,而由顶端细胞分裂形成胚本身。在胚中,又明显表现出极性,即在胚的一端形成胚根,另一端形成胚芽。极性一旦建立,即难于逆转。
植物的再生实验能证明植物体中存在极性。再生(regeneration)是指植物体的离体部分具有恢复植物体其他部分的能力。如植物组织培养中的外植体(包括器官、组织或细胞)都能培育成植株。又如,将柳树枝条悬挂在潮湿的空气中,枝条基部切口附近的一些细胞可能由于受生长素和营养物质的刺激而恢复分生能力,形成愈伤组织,再分化出不定根和芽。无论柳树枝条是正挂还是倒挂,总是在其形态学的上端长芽,形态学的下端长根,而且越靠形态学上端切口处的芽越长,越靠形态学下端切口处的根越长(图8 – 11 )。对根的切段来说,也是形态学的上端长芽,形态学的下端长根。高等植物各种器官的极性强弱不同,一般来说,茎的极性最强,根次之,叶的极性很弱。
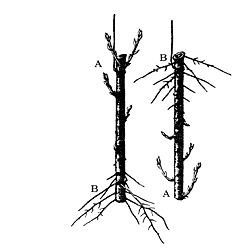
图8–11 柳树枝条的极性与再生
A、形态学上端;B、形态学上端
在植物器官再生过程中,总是形态学的上端长芽,形态学的下端长根,与生长素的极性运输有关。生长素在茎中极性运输,集中于形态学的下端,有利于根的形成,而生长素含量少的形态学上端则发生芽的分化。另外,由于不同器官生长素的极性运输强弱不同,如茎>根>叶,因此不同器官的极性强弱也存在差异。
植物的极性现象在指导生产实践有重要意义。在扦插繁殖时,应注意将形态学的下端插入土中,而不能颠倒。在嫁接时,砧木和接穗要在同一个形态学方向,才能成功。
8.5.5植物生长的相互竞争和相生相克(化感作用)
植物个体的生长要受到与它周围一起生活的植物和其他生物的影响。植物体之间的关系可概括为竞争、共生和寄生这三种关系。① 竞争(competition)是指同种或异种的两个或更多个体,利用共同的有限资源,从而发生对环境资源争夺的现象。绿色植物的竞争主要是对光、水、矿质养分和生存空间的竞争,两个种越相似,对相同资源的竞争也就越激烈。② 共生(symbiosis)关系又分为互惠共生和附生(也称为偏利共生)。互惠共生(mutualism)是指所有有利于共生双方的相互作用,如菌根、根瘤和地衣等现象;附生(epiphytism)是指两个种之间的关系只对一方有利,对另一方无利害的共生。附生植物的产生是长期进化的结果,它们避开了生长在土壤上与其他植物的竞争,使植物能获得更合适的生长条件,如光照。但附生植物过多的繁殖生长,也可引起对宿主不利的影响,甚至导致宿主的死亡,使它们的关系转变为拮抗作用(antagonism)。③ 寄生(parasitism)是指某一物种的个体依靠另一物种个体的营养而生活的现象。寄生于其他植物上并从中获得营养的植物称寄生植物,如菟丝子。寄生植物会使寄主植物的生长减弱,轻者引起寄主植物的生物量降低,重者引起寄主植物的养分耗竭,并使组织破坏而死亡。
植物体也可以通过改变生态环境来影响其他生物体,这表现在两个方面:一方面是它们之间相互竞争(allelospoly),即对环境生长因子,如光照、水分、矿质元素和生存空间等的竞争,由于同种或异种的植物个体利用共同的有限资源,从而发生对环境资源争夺的现象。例如,高秆植物对矮秆植物生长的影响;杂草的滋生蔓延等。另一方面是它们之间相生相克(allelopathy),亦称为化感作用。化感作用的概念是Molish于1937年首次提出的。1984年(E. L. Rice)形成了比较公认概念,即生活的或腐败的植物通过向环境释放化学物质而产生促进或抑制其他植物生长的效应。植物一般通过地上部分茎叶挥发、淋溶和根系分泌物以及植物残株的分解等途径向环境中释放化学物质,从而影响周围植物(受体植物)的生长和发育。植物的化感作用广泛存在于自然界中,与植物间的光照、水肥和空间的竟争一起构成了植物之间的相互作用,它在森林更新、植被演替以及农业生产中具有重要的意义。
对引起化感作用的物质的分离和鉴定已有很多研究,涉及的化学物质很多,它们几乎都是一些分子量较小、结构较简单的植物次生物质,如阿魏酸、对-叔丁基苯甲酸、醚、醛、酮、酚类、有机酸类物质,但许多具体物质并没有分离出来。
植物所产生的化感物质能明显影响植物的种间关系。有些植物产生的化感物质能促进它周围植物的生长,如茄的提取物对水稻生长有促进作用;禾本科与豆科植物(如小麦与豌豆、玉米与大豆)混种,相互有促进作用;在种过苜蓿的土壤里种植番茄、黄瓜、莴苣等植物,生长良好;皂荚、白蜡与七里香,黑果红瑞木与白蜡槭一起生长时,相互之间都有明显的促进作用。研究更多的是一种植物产生的化感物质对另一种植物的抑制作用,如番茄植株释放鞣酸、香子兰酸、水杨酸等能严重抑制莴苣、茄子种子的萌发和幼苗的生长,对玉米、黄瓜、马铃薯等作物的生长也有抑制作用;小麦提取物能抑制繁缕、反枝苋和升马唐等的生长;一些水稻品系能抑制稗和异型莎草的生长;植物残体也会产生化感物质,如玉米、小麦、燕麦和高粱的残株分解产生的咖啡酸、氯原酸、肉桂酸等抑制高粱、大豆、向日葵、烟草生长;小麦残株腐烂产生异丁酸、戊酸和异戊酸,抑制小麦本身生长;水稻秸秆腐烂产生羟基苯甲酸、苯乙醇酸、香豆酸等,抑制水稻秧苗的生长;甘蔗残株腐烂产生羟基苯甲酸、香豆酸、丁香酸等,抑制甘蔗截根苗的萌发与生长。
化感作用的研究,对农林业生产具有重要意义。某些树种不能与另一些树种种在一起,如白桦与松树;苹果树旁不能种玉米等等。在作物布局上可利用有益的作物组合,尽量避免与相克的作物为邻,对有自毒的应避免连作。在农业杂草防除上,还可以通过对化感物质的确定,以便人工合成该物质,不仅可以用来控制杂草,而且因为这种化学物质的易降解性,不会给土壤环境带来污染。
8.6植物的运动
高等植物虽然不能象动物或低等植物那样的整体移动,但是它的某些器官在内外因素的作用下能发生有限的位置变化,这种器官的位置变化就称为植物运动(plant movement)。植物从接受刺激到产生运动一般包括三个基本步骤:①刺激感受(perception ),植物体内的感受器感受环境中的刺激。② 信号传导(transduction ),感受器接受的刺激转换成一种信号,继而把信号传递到产生运动的器官。一般来说,这种信号是对环境刺激产生反应的某种化学或物理的变化,如动作电位、化学信号的激素等。③ 运动反应(motor response)。关于前两个步骤详见第5章(植物的信号转导)。
高等植物的运动可分为向性运动(tropic movement)和感性运动(nastic movement)。
8.6.1向性运动
向性运动是指植物的某些器官由于受到外界环境中单方向的刺激而产生的运动。
根据刺激因素的不同,向性运动可分为向光性、向重性、向化性等。并规定向着刺激方向运动的为正运动,背着刺激方向运动的为负运动。
所有的向性运动都是生长性运动,都是由于生长器官不均等生长所引起的,是不可逆的运动。当器官停止生长或者除去生长部位时,向性运动随即消失。
8.6.1.1向光性
植物生长器官受单方向光照射而引起生长弯曲的现象称为向光性(phototropism)。根据植物向光弯曲的部位不同,可将向光性分为三种类型:⑴ 植物器官向着光照的方向弯曲,称为正向光性(positive phototropism),如向日葵、棉花等植物的茎向光源方向弯曲。⑵ 植物器官背着光照的方向弯曲,称为负向光性(negative phototropism),如拟南芥和常春藤的气生根背光弯曲。⑶ 植物器官保持与光照方向垂直的能力,称为横向光性(diaphototropism),如叶片通过叶柄扭转使其处于对光线适合的位置。
植物感受光刺激的部位主要是嫩茎尖、芽鞘尖端、根尖、某些叶片或生长中的茎等。燕麦、小麦、玉米等禾本科植物的黄化幼苗以及豌豆、向日葵的上下胚轴等,都常用作向光性的研究材料。向光性是植物的一种生态反应,如茎叶的向光性,能使叶子尽量处于吸收光能的最适合位置进行光合作用。
植物向光性是由光照引起的,那么向光性反应的光受体是什么?实验发现,燕麦胚芽鞘对向光性起主要作用的光是420~480nm 的蓝光,其峰值在445nm 左右,其次是360~380 nm紫外光,其峰值在370nm (图8 – 12 )。比较燕麦胚芽鞘向光性的作用光谱和β-胡萝卜素以及核黄素的吸收光谱,发现二者类似,但又不完全相同。因此,有人认为向光性的光受体是与β-胡萝卜素及核黄素类似的物质,有人甚至认为这两种色素就是向光性的光受体, 更多研究者认为可能是黄素蛋白质类。光受体接受光照刺激后,所引起的原初反应,目前仍然不清楚。但光受体接受光照刺激后所引起的一系列反应最终使器官的向光面和背光面不均等生长的原因,有两种对立的看法:一是生长素分布不均匀;二是抑制物质分布不均匀。长期以来,人们认为不均等生长是由于单侧照光后导致生长素在向光面和背光面分布不均匀而引起的。温特Went(1928)用生物测定法证明,燕麦胚芽鞘经单侧照光后,背光一侧顶部扩散到琼脂的生长素活性比向光一侧多,二者的分布比率约为向光面32%,背光面68%(相对比值为27∶57 )。至于照光引起生长素不均等分布的原因,可能是单方向光照引起燕麦胚芽鞘尖端不同部位产生电势差,向光的一侧带负电荷,背光的一侧带正电荷,由于生长素(吲哚乙酸)呈弱酸性带负电荷,因而向带正电荷的背光一侧移动,背光一侧生长素含量升高,生长加快,植株便向光弯曲。
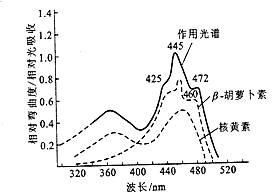
图 8–12 燕麦胚芽鞘的向光性的作用光谱与β-胡萝卜素及核黄素的吸收光谱比较
20世纪80年代以来,许多学者提出向光性的产生是由于抑制物质分布不均匀的看法。他们采用气相-质谱等物理化学方法测得单侧照光后,黄化燕麦芽鞘、绿色向日葵下胚轴和绿色萝卜下胚轴都会向光弯曲,但向光一侧和背光一侧的生长素含量并没有差别(表8 – 7)。相反,却发现向光一侧的生长抑制物质的含量多于背光一侧,而且与光强呈正相关(表8 – 8)。有实验证明,萝卜下胚轴的生长抑制物质可能是萝卜宁(raphanusanin)和萝卜酰胺(raphanusamide),向日葵下胚轴的生长抑制物质可能是黄质醛。燕麦胚芽鞘的生长抑制物质尚不清楚。
表8–7 向日葵、萝卜和燕麦向光性器官中IAA的分布(李合生,2002)
|
器官 | IAA分布(%) |
测定方法 | ||
| 向光一侧 | 背光一侧 | 黑暗(对照) | ||
| 绿色向日葵下胚轴 | 51 | 49 | 48 | 分光荧光法 |
| 绿色萝卜下胚轴 | 51 | 49 | 45 | 电子俘获检测法 |
| 黄化燕麦芽鞘 | 49.5 | 50.5 | 50 | 电子俘获检测法 |
表8 – 8 绿色向日葵下胚轴受单侧光照射后生长抑制剂活性的相对分布(Franssen,1981)
| 抑制剂(%) | |||
| 刺激持续时间(min) | 向光一侧 | 背光一侧 | 弯曲度 |
| 0 | 51.5 | 48.5 | 0° |
| 30~45 | 66.5 | 33.5 | 15.5° |
| 60~80 | 59.5 | 40.5 | 34.5° |
注:采用水芹根生物测定法,幼苗平均抑制物含量相当于每克鲜重60ng顺式黄质醛
据此,有人提出单方向光照导致生长抑制物质在向光一侧积累,胚芽鞘向光一侧的生长受到抑制,因而向光弯曲。生长抑制剂抑制生长的原因可能是妨碍了IAA与IAA受体结合,减少IAA诱导与生长素有关的mRNA的转录和蛋白质的合成。还有实验表明,生长抑制物质能阻止表皮细胞中微管的排列,引起器官的不均衡伸长。
8.6.1.2向重力性
植物在重力的影响下,保持一定方向生长的特性,这种特性称为向重力性( gravitropism )。根顺着重力方向向下生长,称为正向重力性( positive gravitropism );茎背离重力方向向上生长,称为负向重力性( negative gravitropism );地下茎、侧枝、叶柄、次生根等以垂直于重力的方向水平生长,称为横向重力性( diagravitropism )。通常初生根有明显的正向重力性,次生根趋于水平生长;主茎有明显的负向重力性,但侧枝、叶柄、地下茎却偏向水平生长。根的正向重力性有利于根向土壤中生长,以固定植株并吸收水分和矿质。茎的负向重力性则有利于叶片伸展,并从空间上获得充足的空气和阳光。如果将植物幼苗横放,过一段时间后,根会向下弯曲,而茎则向上生长;作物由于风或其他原因倒伏后,茎会弯曲向上生长等现象都是植物向重力性的表现。近年来的太空实验证明,在无重力作用的条件下,植物的根和茎都不发生弯曲。
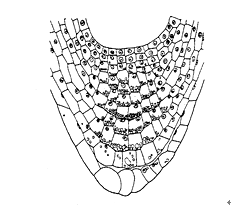
图8–13 两栖焊菜(Roricaamphilia)根冠中的淀粉粒
对植物的向重力性现象长期以来一直用平衡石学说来解释。平衡石(statolith)原指甲壳类动物的一种器官中管理平衡的砂粒,起着平衡的作用。植物的平衡石是一种淀粉体(amyloplast),植物体内平衡石的分布因器官而异。根部的平衡石在根冠的柱细胞中,茎部的平衡石分布在维管束周围的淀粉鞘中。在植物器官变换位置时,确实可以观察到平衡石的移动,并对原生质产生一种压力,这种压力则是被细胞感受的刺激。接受重力刺激的部位是根、茎和胚芽鞘的尖端。有人发现拟南芥突变体不能合成淀粉,但仍有向重力性反应,提出无淀粉突变体中无淀粉质体可能是平衡细胞中的平衡石。
细胞感受到由于平衡石的“沉降”而带来的刺激后,如何导致器官的弯曲生长呢?一般认为,向重力性导致的弯曲生长是由于生长素的不均匀分布引起的。当植株水平放置时,由于重力的作用,器官上侧的生长素将向下侧移动,从而导致上侧生长素减少,下侧生长素增多。生长素向下侧移动可能是由于重力的影响而造成的电势差引起,上侧带负电荷,下侧带正电荷。带负电荷的生长素于是就向下移动。由于根对生长素很敏感,微量的生长素促进根的生长,生长素稍多时,根的生长就受到抑制。因此,根横放时,下侧的生长将由于生长素增多而受到抑制,根就向下弯曲生长,呈正向重力性。与此相反,由于茎对生长素不敏感,因而茎横放时,下侧的生长将由于生长素的增多而加快,茎就向上弯曲生长,呈负向重力性。
根冠中含有脱落酸。图8 – 14说明了ABA影响弯曲,将玉米根冠作各种处理,则得出不同结果。根横置时,根冠内的脱落酸向下移动,然后向根的生长部位(如分生区和伸长区)运输,该处下侧的脱落酸含量较高,于是抑制该侧细胞分裂和伸长,根向下弯曲生长(A、B处理),C、D处理则向上弯曲生长。
近年来的研究表明,Ca2+在向重力性反应中起着重要作用(图8 – 15)。均匀地外施45Ca2+ 于根上,水平放置,发现45 Ca2+向根的下侧移动。将含有钙离子螯合剂的琼脂块放在横放玉米根的根冠上,无重力反应;如改用含Ca2+ 的琼脂块,则恢复向重力性反应。进一步研究发现,玉米根内有钙调素,根冠中的钙调素浓度是伸长区的4倍。外施钙调素的抑制剂于根冠,则根丧失向重力性反应。
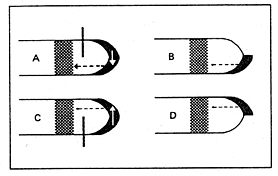
图 8–14 不同处理对玉米根弯曲生长的影响
A、C 生长部位上下方分别插入云母片,将B、D的根冠各切去一半。结果,A、B处理都向下弯曲,A向下弯曲大于B,C、D处理都向上弯曲,C 向上弯曲略小于D
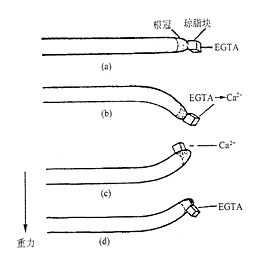
图8–15 Ca2+ 对玉米根向重力性反应的影响(引自Poovaiah,1987)
(a) 涂在根冠上的EDTA妨碍向重力性反应;(b)EDTA预处理后涂上Ca2+恢复向重力性反应;(c)Ca2+涂在根冠一侧引起向Ca2+侧弯曲;(d)EDTA涂在根冠一侧引起偏离EDTA弯曲
综上所述,结合平衡石、生长素、Ca2+ 、钙调素和脱落酸对向重力性的影响,有人提出向重力性的机理:根横放时,平衡石“沉降”到细胞下侧的内质网上,产生压力,诱发内质网释放Ca2+ 到细胞质中,Ca2+ 和钙调素结合,激活细胞下侧的生长素泵和钙泵,引起细胞下侧生长素和Ca2+ 的积累(图8 – 16)。以此同时,在横放根的下侧积累较多的ABA,从而抑制下侧的生长,引起根尖向下弯曲生长。
根的向重力性具有重要的生物学意义。当种子播种到土中,不管胚的方向如何,总是根向下生长,茎向上生长。禾谷类作物倒伏后,茎节可向上弯曲,恢复直立生长。
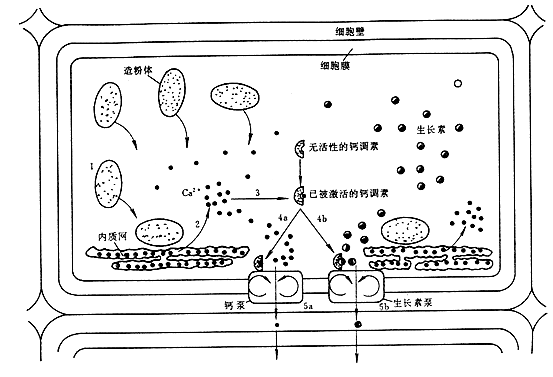
图8–16 向重力性的机理
1、根冠的淀粉体受到重力影响,向下运动压在内质网上;2、诱使内质网将Ca2+释放出来;3、Ca2+与钙调素结合,呈激活状态;4、激活钙泵和生长素泵;5、前者将Ca2+运到细胞壁,后者将生长素运到细胞壁。生长素大部分分布在根的下侧,Ca2+也促进生长素返回伸长区下侧,这样,下侧生长素过多,抑制伸长区伸长,而上侧生长素较少,生长正常。由于上侧生长快,下侧生长慢,所以根就向重力方向弯曲生长。
8.6.1.3 向化性和向水性
向化性(chemotropism)是由于某些化学物质在植物体内外分布不均匀引起的向性生长。 植物根的生长就有向化性。根在土壤中总是朝着肥料多的地方生长。生产上采用深耕施肥,就是为了使根向深处生长,从而可以吸收更多营养。此外,花粉管的生长也表现出向化性,当花粉落在柱头上,凡是亲和的,在胚珠细胞分泌物(如退化助细胞释放的Ca2+ )的诱导下花粉管向胚珠生长。
根的向水性(hydrotropism)也是一种向化性。当土壤干燥而水分分布不均时,根总是趋向潮湿的地方生长,干旱土壤中根系能向土壤深处伸展,其原因是土壤深处的含水量较土表高。种植香蕉、竹子时,可以采用以肥养芽的措施,把它们引到人们希望它生长的地方出芽生长,就是利用根和地下茎在水肥充足的地方生长较为旺盛的生长特点。
8.6.2感性运动
感性运动(nastic movement)是指由没有一定方向的外界刺激所引起的运动,运动的方向与外界刺激无关。根据外界刺激的种类又可分为感夜性、感热性、感震性等。感性运动多数是由细胞膨压的变化所引起,因而也称为紧张性运动(turgor movement)或膨胀性运动,如感震性、感夜性。但也有一些感性运动是由生长的不均匀引起的,如感热性运动。
8.6.2.1感夜性
感夜性运动(nyctinasty movement)主要是由昼夜光暗变化引起的。例如一些豆科植物(如大豆、花生、合欢、木瓜、含羞草、酢浆草等)的叶子,白天叶片张开,夜间合拢或下垂。三叶草、酢浆草的花和蒲公英等许多菊科植物的花序在晚上闭合,白天开放;月亮花、甘薯、烟草等植物花的昼闭夜开,也都是由光引起的感夜性运动。感夜运动的器官是叶柄基部的叶枕或小叶基部,也有一些没有叶枕。在叶枕或小叶基部上下两侧,其细胞的体积、细胞壁的厚薄和细胞间隙的大小都不同,当细胞质膜和液泡膜因感受光的刺激而改变其透性时,两侧细胞的膨压变化也不同,使叶柄或小叶朝一定方向弯曲。感夜运动产生的原因就是由于叶柄基部叶枕的细胞发生周期性的膨压变化所致。叶片在白天合成生长素,运到叶柄下半侧,K+和Cl–也运到生长素浓度高的地方,水分就进入叶枕,细胞膨胀,导致叶片高挺。到晚上,生长素合成和运输量均减少,进行相反过程,叶片下垂。在此过程中,光敏色素在接受光暗变化的刺激中起重要作用,且这种昼夜有内在节奏的变化是由生物钟控制的。
8.6.2.2感热性
由于温度变化而使器官背腹两侧不均匀生长引起的运动,称为感热性运动(thermonasty movement)。如番红花和郁金香花的开放或关闭受温度变化的影响,在温度升高时,花朵开放;温度下降时,花瓣合拢;如将番红花和郁金香从较冷处移至温暖处后,很快又会开花。花的这种感热性运动是不可逆的生长性运动,是由于花瓣上下组织生长速率不同所致。花的感热性运动对植物来说具有重要的意义,因为这样可使植物在适宜的温度下进行授粉,并且可保护花的内部免受不良条件的影响。
8.6.2.3感震性
植物的感震性运动(seismonasty movement)是由于机械刺激而引起的与生长无关的植物运动。含羞草叶片的运动是典型的感震性运动。含羞草不仅在夜晚将小叶合拢,叶柄下垂,即使在白天,当部分小叶受到震动(或其他刺激如烧灼、电刺激)时,小叶也会成对地合拢。如刺激较强,这种刺激可以很快地依次传递到邻近的小叶,甚至可传递到整个复叶的小叶,引起复叶叶柄下垂。如果是更强刺激,则会使整株植物的小叶合拢,复叶叶柄下垂。但经过一定时间,整株植物又可以恢复原状。含羞草对震动的反应很快,刺激后0.1s就开始,几秒钟就完成。上下转递速度极快,可达40~1250px/s。另外,食虫植物捕蝇草的触毛对机械触动产生的捕食运动也是一种反应速度很快的感震性运动。
含羞草叶子感受震动的机理,被认为是由于小叶和复叶叶柄基部叶褥细胞膨压的改变而引起的。叶褥是含羞草叶柄基部的一群特殊细胞,具有特殊的解剖结构。小叶叶褥上半部细胞的间隙较大,细胞壁较薄。当小叶受到震动或其他刺激时,叶褥上半部细胞的透性加大,细胞内的水分和溶质排入细胞间隙,细胞的膨压下降,组织疲软,而此时叶褥下半部的细胞还保持紧张状态,因而小叶成对合拢。一定时间后,水分和溶质又回到叶褥上半部细胞中,小叶又张开。复叶叶褥的结构则正好与小叶叶褥的结构相反,即复叶叶褥的上半部细胞排列紧密,细胞壁较厚,而下半部细胞的间隙较大,细胞壁较薄,受到外界震动或其他刺激导致膨压改变时,复叶就下垂。
震动刺激被植物感受后,转换成什么样的信号,并引起细胞膨压的变化呢?对此,目前有两种不同的看法(参考5.植物的信号转导)。一种认为是由电信号的传递,诱发了感震性运动;另一种认为是由化学信号的传递,诱发了感震性运动。20世纪80年代以来许多学者对此进行了深入的研究。有些学者认为是电传递。震动植株,植株就会产生动作电位(action potential),并有一个特征高峰(图8 – 17)。含羞草的动作电位类似于动物神经细胞,但比较慢。具体过程是,含羞草的小叶和食虫植物捕蝇草的触毛接收刺激后,其中感受刺激细胞的膜透性和膜内外的离子浓度会发生瞬间改变,引起膜电位的变化。感受细胞膜电位的变化还会引起邻近膜电位的变化,从而引起动作电位的传递,当其传至产生动作的部位后,使动作部位细胞膜的透性和膜内外的离子浓度发生改变,从而造成膨压变化,引起感震运动。有些学者认为还存在化学信号传递。目前有人已从含羞草、合欢等植物中分离到一些化合物,它们能引起这些植物的叶褥细胞膨胀,其结构是没食子酸的β-葡萄糖苷。从其他植物中获得这样一些类似物质。有人把这一类物质称为膨压素(turgorins),因为它们能使叶枕细胞膨胀,并且在10-5到10-7M时就有活性。Kallas等人证实,在含羞草质膜外侧有它们专一性的受体存在,推测可能是一种蛋白质。从感震反应的速度来看,似乎动作电位更可能是刺激感受的传递信号。
感震性运动不是由生长的不均匀所引致,而是由细胞膨压的改变而引起的,因而是一种可逆性的运动。
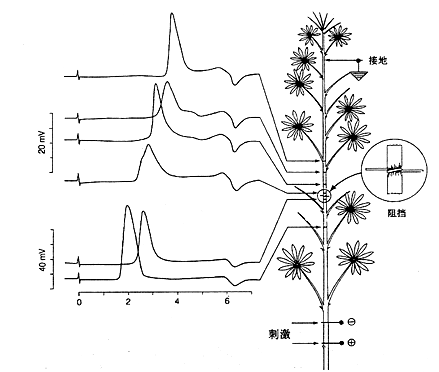
图8-17 测定羽扇豆(lipinus angustifolius)动作电位的实验和结果
8.6.3近似昼夜节奏—生理钟
植物的很多周期性生理活动是环境因素的周期性变化引起的,但也有一些生理活动是由植物体内部的测时系统控制的,不决定于环境条件的变化。如菜豆叶片的昼夜运动(图8 – 18)。菜豆叶片在白天呈水平方向伸展,而在夜间呈下垂状态。即使在外界连续光照或连续黑暗以及恒温条件下仍能在较长的时间中保持原来的周期性变化。在自然条件下,其周期性运动与昼夜的变化是同步的。但是在弱光和20℃的恒定条件下,叶子平举与下垂的周期约为27 h。菜豆的昼夜运动,是一种内源性节奏现象。由于这种生命活动的内源性节奏的周期是在20~28 h之间,接近24 h,因此称为近似昼夜节奏(circadianrhythm),亦称生理钟(physiologicalclock)或生物钟(biologicalclock)。
近似昼夜节奏的现象在生物界中广泛存在,植物、动物和人类都存在生理钟。高等植物中除了上述的菜豆叶片的感夜运动外,还有花朵开放、气孔开闭、蒸腾作用、细胞分裂、膜的透性、胚芽鞘的生长速度等过程中都存在近似昼夜节奏。
关于生理钟的机理,目前还不清楚。生理钟是植物体内部的测时机制。它可以保证一些生理活动按时进行,如菜豆叶片在黎明前就挺起呈水平状态,显然有利于吸收阳光,进行光合作用。
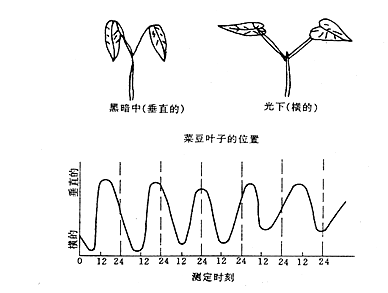
图 8–18 菜豆叶在恒定条件(微弱光,20℃)下的运动
高点代表垂直的叶(左上);低点代表横的叶(右上)
生物节奏的引起必须有一个启动信号。在菜豆叶片的运动中,这个启动信号就是暗期跟随着一个光期,并且具有自调重拨功能。其次,一旦节奏开始,就以大约24 h的节奏自由地运行。植物就是借助于生理钟准确地进行测时过程。
关键词
种子萌发(seedgermination) 胚胎发育晚期丰富蛋白(late embryogenesisabundant protein ,LEA)种子活力(seed vigor)种子生活力(seed viability) 种子寿命(seedlongevity) 顽拗性种子(recalcitrant seed ) 正常性种子(orthodox seed ) 人工种子(artificial seed) 贮存mRNA(long lived mRNA) 植物生长(plant growth) 分化(differentiation) 脱分化(dedifferentiation) 再分化(redefferentiation) 细胞周期(cell cycle) 细胞全能性(totipotency) 组织培养(tissue culture) 外植体(explant) 愈伤组织(callus) 生长大周期(grand period of growth) 伸展蛋白(extensins) 生长的温周期性(thermoperiodicity of growth) 生长的季节周期(seasonal periodicity ofgrowth) 生长的相关性(correlation of growth) 根冠比(root/top ratio, R/T比) 顶端优势(apical dominance或terminal dorminance) 极性(polarity) 再生(regeneration) 化感作用(allelopathy) 植物运动(plantmovement) 向性运动(tropic movement) 向光性(phototropism) 向重力性(gravitropism ) 向化性 向水性 感性运动(nastic movement) 感夜性(chemotropism) 感热性(thermonastymovement) 感震性(seismonasty movement) 近似昼夜节奏(circadian rhythm)
思考题
1.种子的生活力和活力有何不同?用哪个概念更能准确地描述种子萌发的情况?
2.试述种子萌发的三个阶段,以及各阶段的代谢特点。
3.种子萌发过程中吸水的动力是如何变化的?
4.种子萌发的早期能动用胚乳或子叶中的贮藏物质作为呼吸作用的底物吗?长命mRNA是在何时被合成?何时起作用的?
5.总结种子萌发过程中贮藏物质的动员及再利用情况。
6.以根尖为例,说明处在细胞分裂、细胞伸长阶段的细胞以及已分化的细胞在形态结构和生理生化方面有何不同?细胞的分化受哪些因素调控?
7.利用植物组织培养技术将菊花叶的切片培养为一株完整的植株,要经过哪些步骤?
8.用你所学的知识解释“根深叶茂”、“本固枝荣”、“旱长根、水长苗”。
9.产生顶端优势的可能原因是什么?举出实践中利用或抑制顶端优势的2~3个例子。
10.植物地上部与地下部相关性表现在哪些方面?在生产上如何调节植物的根冠比以提高产量?
11.植物生长的最适温度和协调最适温度有何不同?温度“三基点”对生产实践有何指导
意义?
12.试述光对植物生长的影响。
13.简述植物向光性、向重力性的机理。
14.向性运动的类型及产生的原因是什么?
主要参考文献
1.Buchanan B B. Gruissem W. and R L. Jones. 2000. Biochemistry & Molecular Biology of Plants. The American Society of Plant Physiologists, Rockville Maryland.
2、瑞炽主编 植物生理学 北京:高等教育出版社 2001年
3、王忠主编 植物生理学 北京:中国农业出版社 2000年
4、杨世杰主编 植物生物学北京:科学出版社 2000年
5、李合生主编 现代植物生理学北京:高等教育出版社 2002年
6、汪堃仁,薛绍白,柳惠图主编 细胞生物学 北京:北京师范大学出版社 1998年
7、曾广文,蒋德安主编 植物生理学北京:中国农业科技出版社 2000年
8、陈润政,黄上志,宋松泉,傅家瑞主编 植物生理学广州:中山大学出版社,1998年
9、崔澂,桂耀林主编 经济植物的组织培养与快速繁殖 北京:中国农业出版社 1985年
10、 刘承宪 重力植物生理学. 见:余叔文,汤章城主编 植物生理与分子生物学(第二版)北京:科学出版社 1998 . 824~836
11、朱治平种子的形成与萌发. 见:余叔文,汤章城主编 植物生理与分子生物学(第二
版)北京:科学出版社 1998 . 594~609
12、余叔文,孙文浩 植物的它感作用. 见:余叔文,汤章城主编 植物生理与分子生物学(第二版)北京:科学出版社 1998 . 699~720
13、颜昌敬主编 植物组织培养手册 上海:上海科学技术出版社 1990年
14、郑光植编著 植物细胞培养及其次级代谢 云南:云南大学出版社 1992年
15、李浚明编译 植物组织培养教程 北京:中国农业大学出版社 1992年
16、罗士韦,许智宏主编 经济植物组织培养 北京:科学出版社 1988年

