
-
1 课内学习
-
2 课外阅读
第三节 甘油三脂代谢(2h)
一、TG的分解代谢
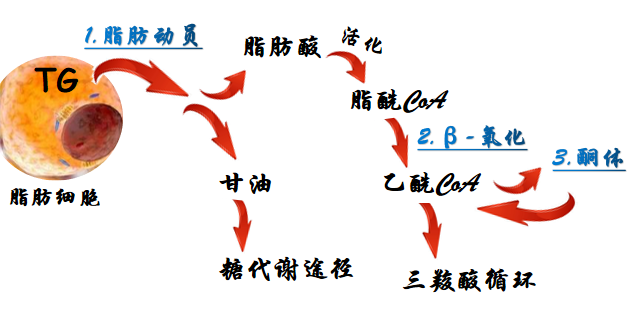
(一)脂肪动员(fat mobilization)
1.概念:储存在脂肪细胞中的脂肪,被脂肪酶逐步水解为游离脂酸及甘油,并释放入血以供其他组织氧化利用的过程。
2.过程:
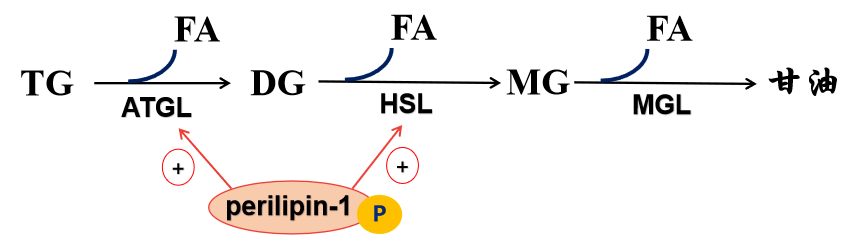
3.关键酶及调节:
(1)脂肪甘油三酯脂肪酶(adipose triglyceride lipase,ATGL)
(2)激素敏感性甘油三酯脂肪酶(hormone-senstive triglyceride lipase,HSL )
(3)脂滴包被蛋白-1(perilipin-1)
(4)脂解激素和抗脂解激素的调节作用。
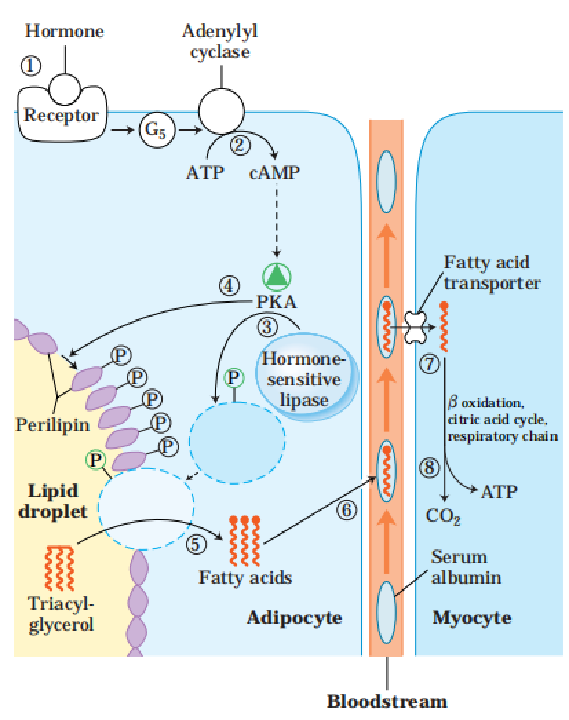
4.产物去向(甘油和脂肪酸)
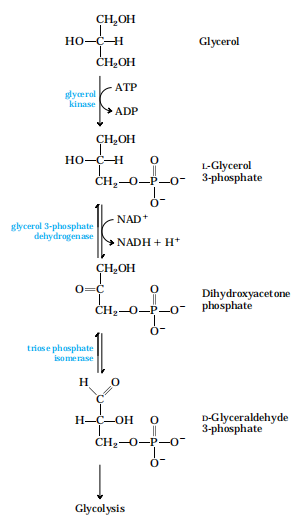
(1)甘油如何利用?进入糖代谢途径
(2)脂肪酸如何利用?经β-氧化生成乙酰CoA进入三羧酸循环,彻底氧化分解为,并释放出大量能量。
(二)脂肪酸的β-氧化
1.部位:动物细胞脂肪酸氧化酶位于线粒体内。肝、心肌、骨骼肌最活跃。RBC、脑组织不能进行FA的氧化。
2.步骤:
(1)脂酸的活化----脂酰CoA的生成
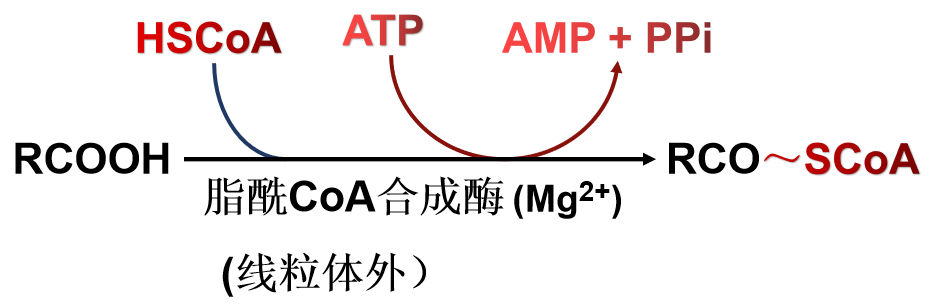
(2)脂酰CoA进入线粒体
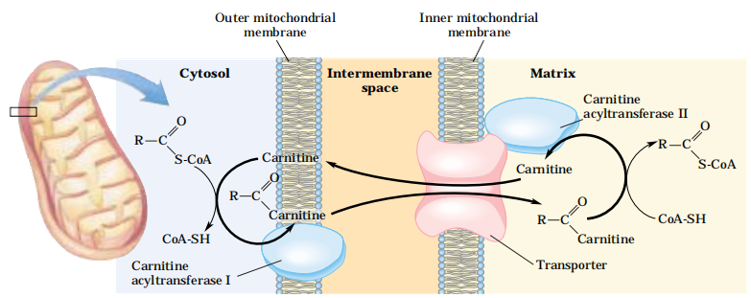
脂酰CoA进入线粒体是脂酸β-氧化的主要限速步骤,如饥饿时,糖供不足,此酶活性增强,脂肪酸氧化增强,机体靠脂肪酸来供能。反之,饱食时脂肪酸合成增强,脂肪酸合成的第一个中间产物丙二酰-CoA抑制肉碱脂酰转移酶1的活性,抑制脂肪酸的氧化。
(3)脂酸β-氧化:在脂酸氧化酶系的顺序催化下,从脂酰基β-C开始,进行脱氢、加水、再脱氢及硫解四步反应,完成一次β-氧化。长链脂肪酸经β-氧化,以乙酰CoA的形式,相继氧化去除2C基团。
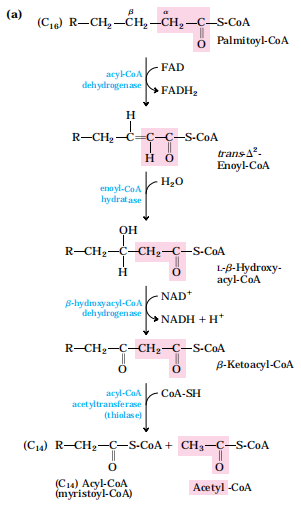
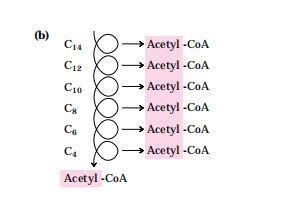
(4)乙酰CoA进入三羧酸循环及氧化磷酸化
3.能量的生成 以16C软脂酸为例
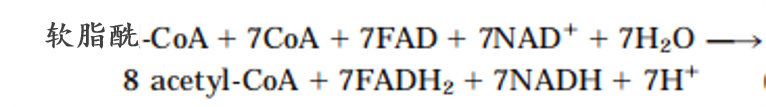
7×2.5+7×1.5+8×10-2=106分子ATP
4.不同的脂肪酸有不同的氧化方式
(三)酮体的生成与利用
概念:β-羟丁酸、乙酰乙酸和丙酮三者统称为酮体,是脂酸在肝分解氧化时特有的中间代谢物。
酮体的生成:在肝细胞线粒体,利用脂肪酸β-氧化生成的乙酰CoA合成
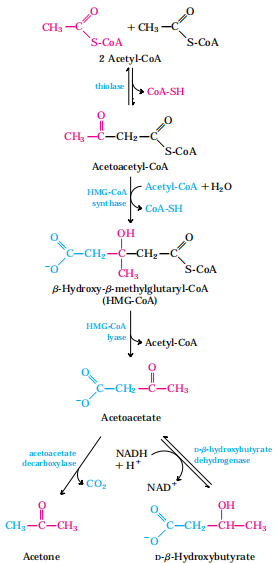
2.酮体的利用:肝外组织有酮体利用酶系
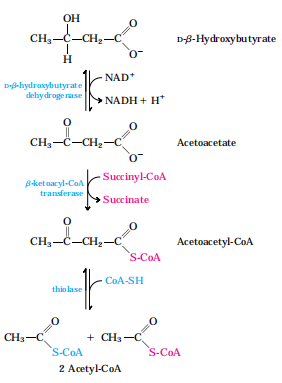
3.酮体生成的意义
酮体在肝内生成,在肝外利用,是肝向外输出能源的一种形式。
(1)易溶于水,便于运输;而且分子小,易通过血脑屏障、毛细血管壁,乙酰乙酸和β-羟丁酸通过血液输送到肝脏以外的组织(肝外组织),它们被转化为乙酰辅酶A,在柠檬酸循环中被氧化,提供了骨骼、心肌和肾皮质等组织所需的大部分能量。当葡萄糖缺乏时,优先使用葡萄糖作为燃料的大脑能够适应在饥饿条件下使用乙酰乙酸或β-羟丁酸。
(2)健康、营养良好的人以相对较低的速度产生酮体,血酮体仅为0.03~0.5mmol/L。
在饥饿期间,糖异生会耗尽柠檬酸循环中间体如草酰乙酸,将乙酰辅酶A转移到酮体的生产中;
未经治疗的糖尿病患者会产生大量的乙酰乙酸,会导致酮症酸中毒,他们的血液中含有大量的丙酮,这是有毒的。 丙酮具有挥发性,使呼吸具有特殊的“烂苹果”气味,这有时对诊断糖尿病是有用的。
4.酮体生成的调节
(1)进食:饱食时胰岛素分泌增加,脂肪动员减少,酮体生成减少;饥饿时,脂解激素分泌增多,脂肪动员增强,脂肪酸氧化加速,酮体生成增加。
(2)糖代谢:旺盛,3-磷酸甘油及ATP充足,脂肪酸脂化增多,氧化减少,酮体生成减少;代谢障碍,乙酰辅酶A进入TCA受阻而堆积,产生酮体增加。
(3)丙二酰CoA:糖代谢过程中的乙酰CoA和柠檬酸能别构激活乙酰CoA羧化酶,促进丙二酰CoA合成,而后者能抑制肉碱脂酰转移酶Ⅰ,阻止β-氧化的进行,酮体生成减少。
二、TG的合成代谢
(一)合成场所(内质网胞液侧)
肝,合成能量最强,但不能储存。以VLDL(极低密度脂蛋白)的形式运输到脂肪组织储存。
脂肪组织,既能合成也能储存,是机体TG的储库。
小肠,利用食物消化吸收的脂类物质,再次合成TG。以CM的形式运输的脂肪组织储存。(详见消化吸收)
(二)合成原料(2种组成成分的活化)
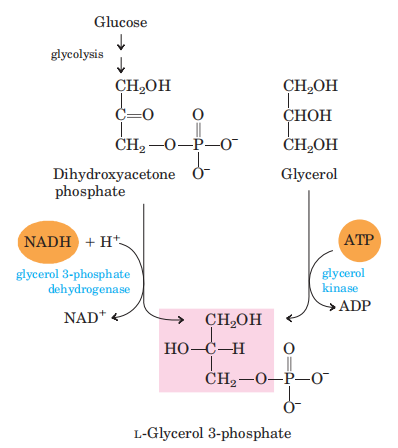
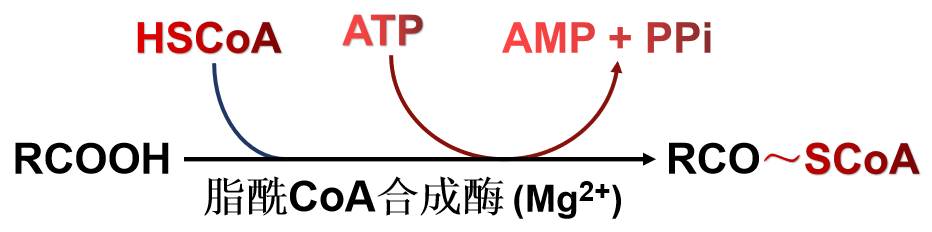
(三)合成途径
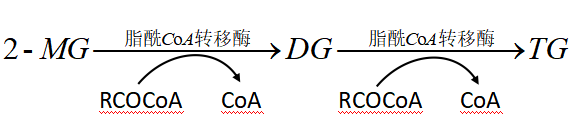
1.甘油一酯途径(小肠粘膜细胞)
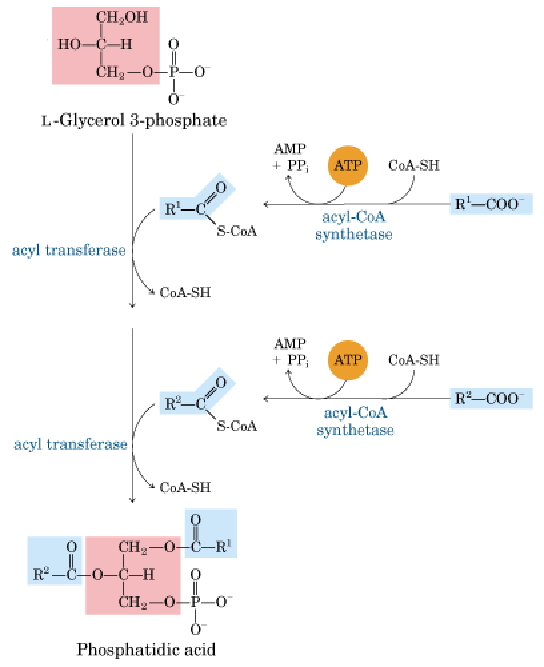
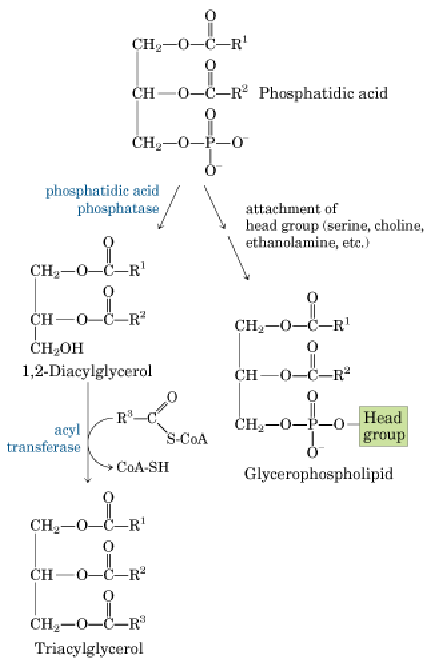
2.甘油二酯途径(肝、脂肪细胞)
三、脂肪酸的合成
(一)软脂酸的合成
1.部位:脂肪酸合成酶系在胞液中,肝, 肾, 脑, 肺, 乳腺, 脂肪等多种组织可以合成脂肪酸,肝细胞内的酶活性最高。
2.原料:
(1)C源,乙酰CoA,来源于糖代谢,在线粒体内生成,需要经过柠檬酸-丙酮酸循环转运到胞液中。
(2)H源,NADPH,主要来源磷酸戊糖途径,少部分可由柠檬酸-丙酮酸循环提供。
(3)能源,ATP;还有Mn2+、HCO3-等
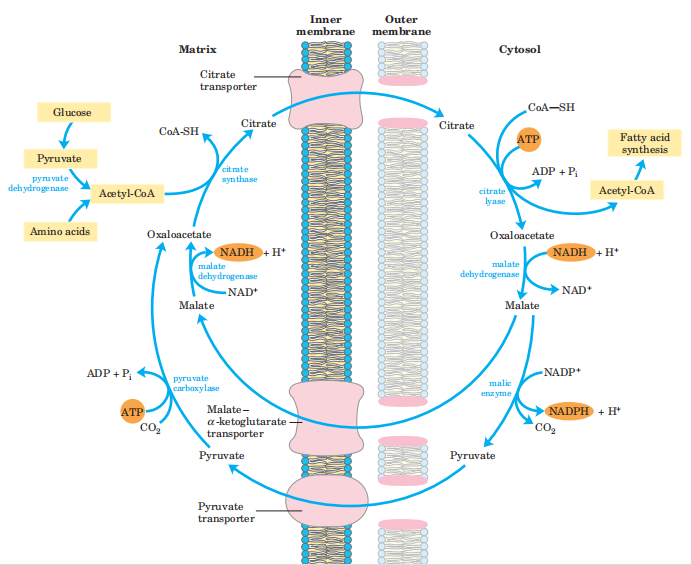
3.合成过程:
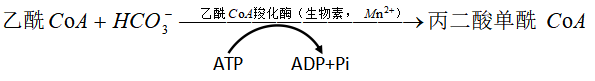
(1)丙二酰CoA的合成,限速步骤,催化反应的酶是乙酰辅酶A羧化酶(生物素),是调节脂肪酸的重要部位。
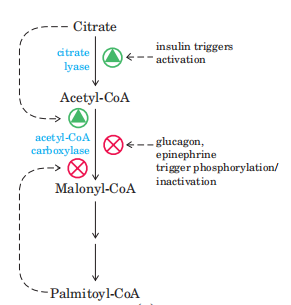

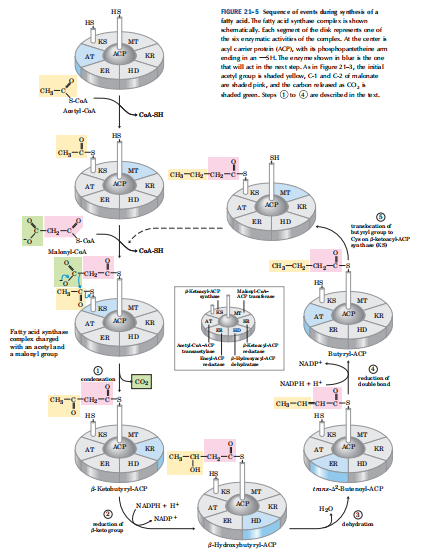
(2)软脂酸的合成,脂肪酸合酶催化脂肪酸碳链以重复的四步序列组装(缩合、加氢、脱水、再加氢),随着每一轮循环,脂酰链延伸2个C。当链长度达到16个C时,产物(软脂酸,16:0)离开循环。
表:大肠杆菌脂肪酸合酶复合体的蛋白质
| 成分 | 功能 |
酰基载体蛋白(ACP) | 用硫酯键携带酰基 |
乙酰CoA-ACP转酰基酶(AT) | 酰基从CoA转移到KS的Cys残基 |
β-酮脂酰-ACP合酶(KS) | 缩合酰基和丙二酰基 (KS至少有三个同工酶) |
丙二酰CoA-ACP转移酶(MT) | 将丙二酰基从CoA转至ACP |
β-酮脂酰-ACP还原酶(KR) | β-酮基还原为β-羟基 |
| β-羟脂酰-ACP脱水酶(HD) | 将H2O从β-羟脂酰-ACP中移除, 形成双键 |
烯脂酰-ACP还原酶(ER) | 还原双键,形成饱和酰基-ACP |
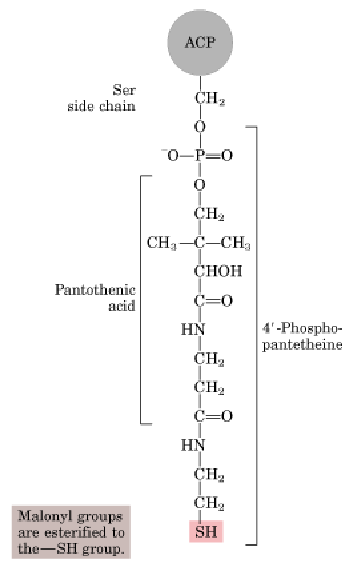
ACP,酰基载体蛋白,辅基为4’-磷酸泛酰巯基乙胺
,
在哺乳动物中,一个单一的大多肽(Mr 240,000)包含所有七种酶活性,还有一种水解活性(它能从酶复合体的ACP部分释放出合成好的脂肪酸)。 有功能的哺乳动物脂肪酸合酶是一个二聚体(Mr 480,000),由两个相同的亚基“头对尾”躺着。形成3个结构域:①AT-MT-KS-SH,催化底物的进入和缩合;②KR-HD-ER-ACP,催化还原反应;③TE,释放出合成好的脂肪酸
(二)软脂酸的延长
1.内质网脂酸碳链延长酶系,以丙二酰CoA为二碳单位供体,可延长至C24,以硬脂酸(18C)为主。
2.线粒体延长酶系,以乙酰CoA为二碳单位供体,可延长至C24~26,以硬脂酸(18C)为主。
(三)不饱和脂肪酸的合成
内质网有脂酰CoA去饱和酶,哺乳动物缺乏Δ9位以上的去饱和酶,只能合成单不饱和脂酸,不能合成多不饱和脂酸,亚油酸(18:2,Δ9,12)、亚麻酸(18:3,Δ9,12,15)、花生四烯酸(20:4,Δ5,8,11,14),必须从食物(主要是植物油)中摄取。
(四)调节
1. 代谢物的调节作用:一方面代谢物充当别构效应剂;另一方面作为合成原料影响供应量
2. 激素的调节作用:胰岛素和胰高血糖素的作用
重点内容:脂肪动员的概念和限速酶;脂肪酸β-氧化的概念,过程及能量的计算;酮体的概念,代谢特点及生理意义;
难点内容:对照脂肪酸的氧化学习脂肪酸合成
本节新进展:脂肪酸合酶抑制剂与抗肿瘤


