
-
1 内容
-
2 测验14.3
一、教学目标
1.掌握DNA的复制过程以及参与DNA复制的一些酶和蛋白质;
2.真核生物与原核生物DNA复制的主要差异;
3.逆转录的过程及其生物学意义。
二、教学重点
1. DNA合成的两条途径;
2. DNA的损伤和修复。
三、教学难点
比较分析DNA复制与逆转录的异同
DNA复制过程(E.coli.)
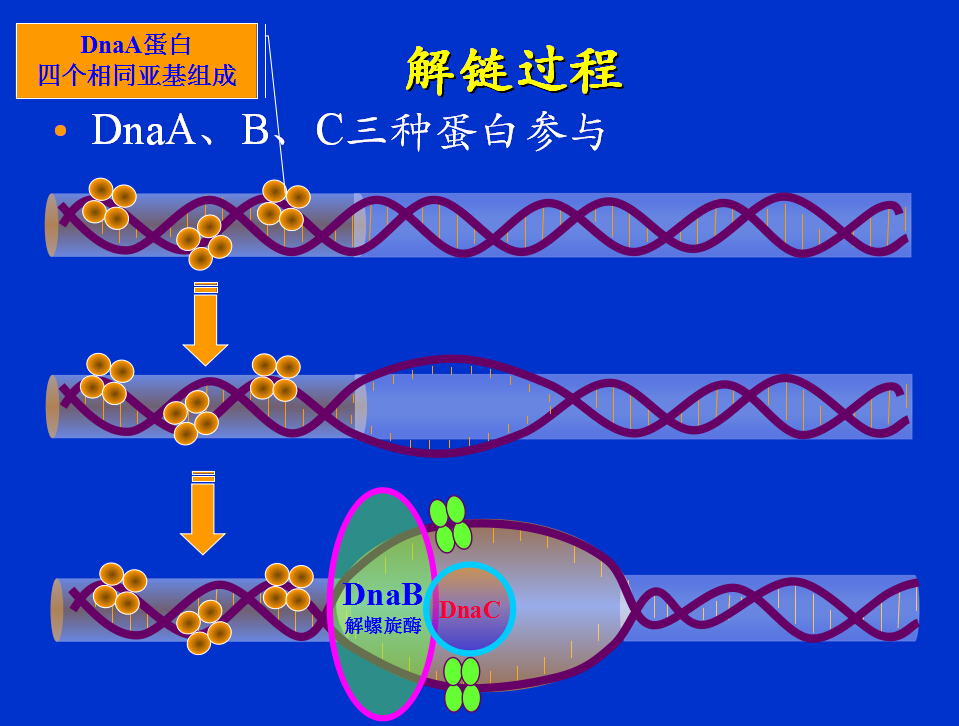
1、复制的起始
引发:当DNA的双螺旋解开后,合成RNA引物的过程。
引发体:引物合成酶与各种蛋白质因子(dnaB、dnaC、n、n'n''I)构成的复合体,负责RNA引物的合成。
引发体沿着模板链5’→3’方向移动(与冈崎片段合成的方向正好相反,而与复制叉移动的方向相同),移到一定位置上即可引发RNA引物的合成。
E.coli.DNA复制原点ori C,由245bp组成,三组13bp重复序列(近5,端处),四组9 bp重复序列(另一端处)。
大肠杆菌复制原点起始复制所需蛋白质:
DNaA 在原点处打开双螺旋 DNaB 使DNA解旋 DNaC DNaB结合在原点所需 Hu 刺激起始 引物酶(DNaG) 合成RNA引物 SSB 结合单链DNA RNA聚合酶 促进DNaA活性 旋转酶 松驰DNA扭曲应力 20个DnaA结合在四组9bp重复区,形成起始复合物,DNA环绕此复合物。
三组13bp重复区依次变性,产生开放型复合物。
DnaB(在DnaC协助下)与开放复合物结合,进一步解链。
2、DNA链的延长反应
前导链只需要一个RNA引物,后随链的每一个冈崎片段都需要一个RNA引物,链的延长反应由DNA pol.Ⅲ催化。
复制体:在DNA合成的生长点(既复制叉上)分布着许多与复制有关的酶和辅助因子,它们在DNA的模板链形成离散的复合物,彼此配合进行高度精确的复制,称为复制体。
复制体沿着复制叉方向前进就合成DNA。
3、RNA引物的切除及缺口补齐
DNA polⅠ的5, → 3,外切活力,切除RNA引物。
DNApolⅠ的5, → 3,合成活性补齐缺口。
4、DNA切口的连接
DNA ligase,动物、真核由ATP供能,原核由NAD供能。
5、DNA合成的终止
环状DNA、线性DNA,复制叉相遇即终止。
小结:
⑴ DNA解螺旋酶解开双链DNA。
⑵ SSB结合于DNA单链。
⑶ DNA旋转酶引入负超螺旋,消除复制叉前进时带来的扭曲张力。
⑷ DNA引物酶(在引发体中)合成RNA引物。
⑸ DNA pol.Ⅲ在两条新生链上合成DNA。
⑹ DNA polⅠ切除RNA引物,并补上DNA。
⑺ DNA ligase连接一个冈崎片段。
DNA复制过程中,聚合酶对dTTP和dUTP的分辨能力高,有少量dUTP掺入DNA链中,此时,U-糖苷酶、AP内切酶、DNA polⅠ、DNA ligase共同作用,切除尿嘧啶,接上正确的碱基。
六、DNA复制的真实性
生物体DNA复制具有高度真实性,复制107-1011碱基对,只有一个错误碱基。
碱基对的自由能通常在4-13KJ/mol,这样的自由能相当于平均参入100个核苷酸就可能出现一次错配,仅靠Watson-Crick双螺旋的碱基配对原则,突变率将高达10-2 。
1、DNA聚合酶对碱基的选择作用
酶的被动论:不同的核苷酸在聚合位点停留时间不同,正确的dNTP能长时间停留,而参与聚合。DNA聚合酶能依照模板的核苷酸,选择正确的dNTP掺入引物末端。
酶积极参与理论:DNA聚合酶对正确与错误的核苷酸,不仅亲和性不同,而且将它们插入DNA引物端的速度也不同。
动力学校正阅读:在新的磷酸二酯键未形成时,dNTP结合在酶与模板—引物复合物的聚合位点上,DNA聚合酶能识别正确与错误的dNTP。
DNA聚合酶对底物的识别作用,DNA聚合酶有两种底物,一种是DNA模板—引物,另一种是dNTP。
DNA聚合酶先识别DNA模板和引物的3,未端,再识别底物dNTP,是一种有序的识别过程。
2、3,→5,外切活性的校正阅读
E. coli. DNA pol.Ⅰ和pol.Ⅲ有3,→5,外切活性,可删除错误插入的核苷酸。
缺失3, →5,外切活性的E. coli. DNA pol.Ⅰ,催化DNA合成时,出现错误的几率增高5-50倍。因此,3,→5,外切活性可以使DNA复制的真实性,提高1-2个数量级。
3、影响DNA合成真实性的因素
⑴高浓度NMP(如3,-AMP, 5,-GMP)
NMP竞争酶的dNTP结合位点,抑制3,→5,外切活性。
⑵某一种dNTP浓度银高,可使引物3,末端离开外切活性中心。
⑶dNTP 一般与二价阳离子结合成活化形式,Mg2+为主要的二价阳离子。当用其它二价阳离子(如Mn2+)代替Mg2+时,会改变酶的主体结构,影响聚合活性和3,→3,外切活性。
4、为什么用RNA引物
⑴从模板复制最初几个核酸时,碱基堆集力和氢键都较弱,易发生错配
⑵新复制的最初几个核苷酸,没有与模板形成稳定双链,DNA聚合酶的5,→3,校对功能难发挥作用。

