
-
1 内容
-
2 测验5.3
一、教学目标
1.了解核苷酸的化学结构及化学性质;
2.掌握核酸二、三级结构及其碱基配对规律;
3.分析比较核酸分子的组成和结构上的特点;
4.联系实际理解核酸性质和生物学功能之间的关系。
二、教学重点
1.碱基、核苷酸的结构、性质
2.核酸的结构、性质和生物功能。
三、教学难点
1 .DNA的结构
2 .RNA的结构
3 .Sanger法测序
DNA的空间结构
一、DNA的二级结构

1953年,Watson和Crick根据Chargaff 规律和DNA Na盐纤维的X光衍射数据提出了DNA的双螺旋结构模型。
1、Watson-Crick双螺旋结构建立的根据
①Chargaff 规律 1950年
a. 所有DNA中,A=T,G=C 且A+G=C+T。
b. DNA的碱基组成具有种的特异性,即不同生物的DNA皆有自己独特的碱基组成。
c. DNA碱基组成没有组织和器官的特异性。
d. 年龄、营养状况、环境等因素不影响DNA的碱基组成。
② DNA的Na盐纤维和DNA晶体的X光衍射分析。
2、Watson-Crick双螺旋结构模型
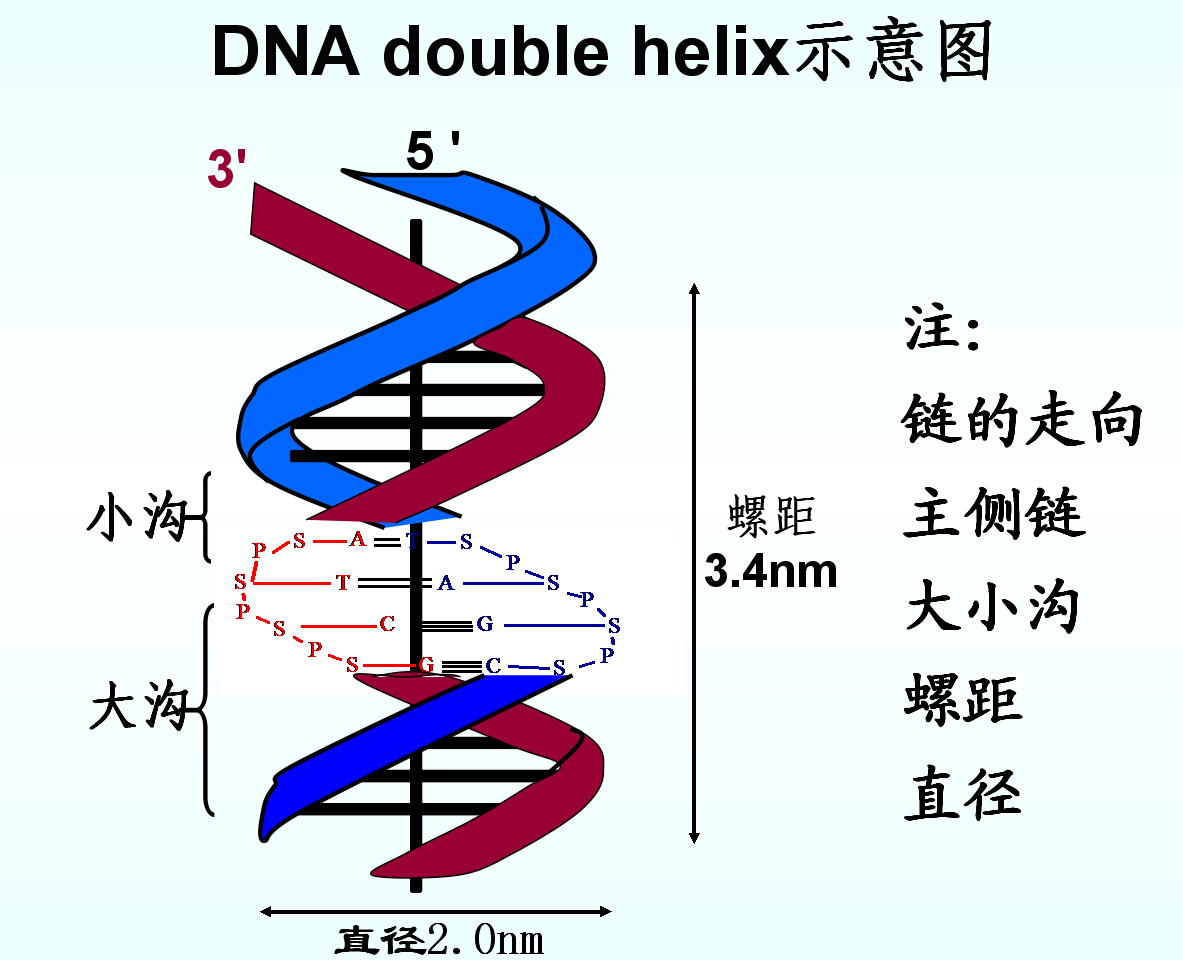
a.两条反平行的多核苷酸链绕同一中心轴相缠绕,形成右手双股螺旋,一条5’→3’,另一条3’→5’
b.嘌呤与嘧啶碱位于双螺旋的内侧,磷酸与脱氧核糖在外侧。磷酸与脱氧核糖彼此通过3’、5’-磷酸二酯键相连接,构成DNA分子的骨架。
c.螺旋平均直径2nm:每圈螺旋含10个核苷酸、碱基堆积距离:0.34nm、螺距:3.4nm
d.两条核苷酸链,依靠彼此碱基间形成的氢链结合在一起。碱基平面垂直于螺旋轴。A=T、G=C
碱基互补原则具有极重要的生物学意义,DNA的复制、转录、反转录等的分子基础都是碱基互补。
3、稳定双螺旋结构的因素
①碱基堆积力(主要因素) 形成疏水环境。
②碱基配对的氢键。GC含量越多,越稳定。
③磷酸基上的负电荷与介质中的阳离子或组蛋白的正离子之间形成离子键,中和了磷酸基上的负电荷间的斥力,有助于DNA稳定。
④碱基处于双螺旋内部的疏水环境中,可免受水溶性活性小分子的攻击。
二、DNA二级结构的不均一性和多型性
(一)DNA二级结构的不均一性
1、反向重复序列(回文序列)
DNA序列中,以某一中心区域为对称轴,其两侧的碱基对顺序正读和反读都相同,即对称轴一侧的片段旋转180°后,与另一侧片段对称重复。
较长的回文结构,在某些因素作用下,可形成茎环式的十字结构和发夹结构。功能还不完全清楚,但转录的终止作用与回文结构有关。
较短的回文序列,可作为一种特别信号,如限制性核酸内切酶的识别位点。
2、富含A T的序列
高等生物中,A+T与C+G的含量差不多相等,但在它们的染色体的某一区域,A T含量可能很高。
在很多有重要调节功能(不是蛋白质编码区)的DNA区段都富含A T碱基对。特别是在复制起点和启动的Pribnow框的DNA区中,富含A T对。这对于复制和转录的起始十分重要,因为G C对有三个氢键,而A T对只有两个氢键,此处双键易解开。
(二)DNA二级结构的多型性
1、B—DNA:典型的Watson-Crick双螺旋DNA
2、A-DNA
3、Z-DNA:左手螺旋的DNA。天然B-DNA的局部区域可以形成Z-DNA。
4、DNA三股螺旋
在多聚嘧啶和多聚嘌呤组成的DNA螺旋区段,序列中有较长的镜像重复时,可形成局部三股螺旋,称H-DNA。
DNA的三链结构常出现在DNA复制、重组、转录的起始或调节位点,第三股链的存在可能使一些调控蛋白或RNA聚合酶等难以与该区段结合,从而阻遏有关遗传信息的表达。
三、环状DNA
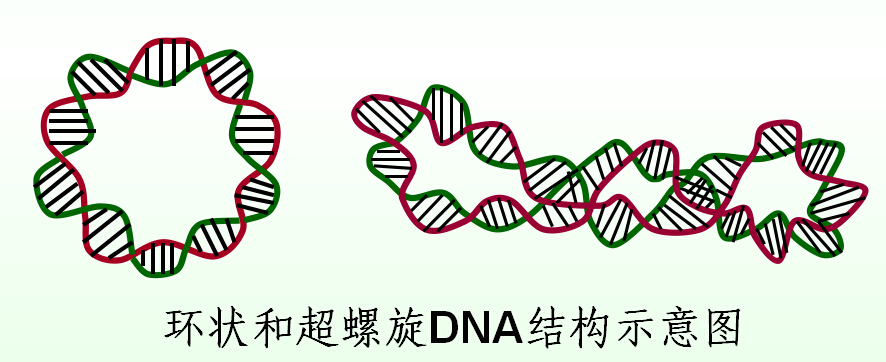
生物体内有些DNA是以双链环状DNA的形式存在,包括:
某些病毒DNA、某些噬菌体DNA、某些细菌染色体DNA、细菌质粒DNA
真核细胞中的线粒体DNA、叶绿体DNA
1、环形DNA的不同构象
(1)松弛环形DNA
线形DNA直接环化
(2)解链环形DNA
线形DNA拧松后再环化
(3)正超螺旋与负超螺旋DNA
线形DNA拧紧或拧松后再环化,成为超螺旋结构。
绳子的两股以右旋方向缠绕,如果在一端使绳子向缠紧的方向旋转,再将绳子两端连接起来,会产生一个左旋的超螺旋,以解除外加的旋转造成的胁变,这样的超螺旋叫正超螺旋。
如果在绳子一端向松缠方向旋转,再将绳子两端连接起来,会产生一个右旋的超螺旋,以解除外加的旋转所造成的胁变,这样的超螺旋称负超螺旋。
2、环形DNA的拓扑学特性
以260bp组成的线形B-DNA为例,螺旋周数260/10.4=25。
①连环数(L)
DNA双螺旋中,一条链以右手螺旋绕另一条链缠绕的次数,以L表示。
松驰环:L=25
解链环:L=23
超螺旋:L=23
②缠绕数(T)
DNA分子中的Watson-Crick螺旋数目,以T表示
松驰环T=25
解链环T=23
超螺旋T=25
③超螺旋周数(扭曲数W)
松驰环W=0
解链环W=0
超螺旋W= -2
L=T+W
④比连环差(λ)
表示DNA的超螺旋程度
λ=(L—L0)/L0
L0是指松驰环形DNA的L值
天然DNA的超螺旋密度一般为-0.03~-0.09,平均每100bp上有3-9个负超螺旋。
负超螺旋DNA是由于两条链的缠绕不足引起,很易解链,易于参加DNA的复制、重组和转录等需要将两条链分开才能进行的反应。
3、拓扑异构酶
此酶能改变DNA拓扑异构体的L值。
①拓扑异构酶酶I(拧紧)
能使双链负超螺旋DNA转变成松驰形环状DNA,每一次作用可使L值增加1,同时,使松驰环状DNA转变成正超螺旋。
②拓扑异构酶酶II(拧松)
能使松驰环状DNA转变成负超螺旋形DNA,每次催化使L减少2,同时能使正超螺旋转变成松驰DNA。
四、染色体的结构
1、大肠杆菌染色体
大肠杆菌染色体是由4.2×106bp组成的双链环状DNA分子,约3000个基因。
这些DNA结合蛋白,使4.2×106bp的E.coli染色体DNA压缩成为一个手脚架形结构,结构中心是多种DNA结合蛋白,DNA双螺旋分子有许多位点与这些蛋白结合,形成约100个小区,每个小区的DNA都是负超螺旋,一个小区的DNA有两个端点被蛋白质固定,每个小区相对独立。
2、真核生物染色体
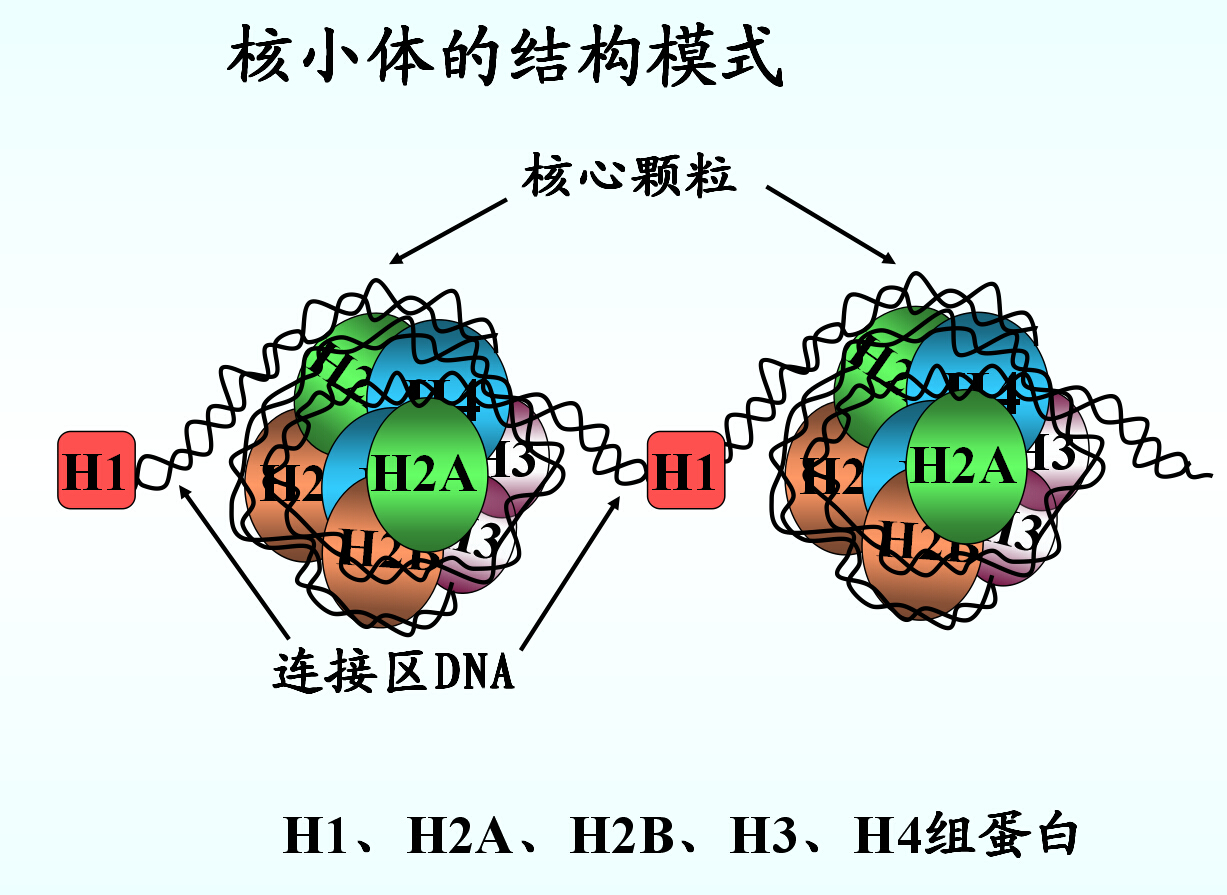
主要由组蛋白和DNA组成。
组蛋白是富含碱性a.a(Lys、Arg)的碱性蛋白质,根据Lys/Arg比值不同,可分为H1、H2A、H2B、H3、H4五种,均为单链蛋白质,分子量11000-21000。
H2A、H2B、H3、H4各两分子对称聚集成组蛋白八聚体。
146bp长度的DNA双螺旋盘绕在八聚体上形成核小体。
核小体间DNA长度15-100bp(一般60bp)其上结合有H1
2H2A、2H2B、2H3、2H4组蛋白八聚体 146bpDNA 核小体 串联 染色质 折叠 染色体
DNA(直径2nm)
盘绕组蛋白八聚体上,结合H1,压缩比1/7
核小体(一级结构)
螺旋化,压缩比1/6
螺线管(二级结构)
再螺旋化,压缩比1/40
超螺线管(三级结构)
折叠,压缩比1/5
染色单体(四级结构)
总压缩比:1/8400~1/10000
五、DNA的生物学功能
首次直接证明DNA的遗传功能的是Avery的肺炎双球菌转化实验。

