

第八章 性别控制
第26课时 水产动物的性别决定与性分化
一、性别控制的意义(理解)
1.单性养殖可提高群体生长率
罗非鱼雄性的生长速度比雌性快;
鲤、鲫、草鱼、鳗鲡等雄性的生长速度比雌性慢;
对虾雌性的体长及体重明显大于雄性;
罗氏沼虾则是雄性大于雌性。
2.控制繁殖速度
全雄控制技术:罗非鱼性成熟早、繁殖快,一年多次产卵,不同世代混养,养殖过程中密度变化过大,抑制整个群体的生长,降低产品质量和养殖效益。罗非鱼的全雄控制技术可有效解决这一问题。
3.延长有效生长期
冷水性的虹鳟,雄性2龄成熟,雌性则在3龄。成熟后无论雌雄,都会出现生长率降低、死亡率提高、产品肉质和外观品质下降的现象。因此,虹鳟一般饲养2年上市。但对雌鱼来说,此时并没有充分长大,而单留雌性养殖又需要花费较多的人力。
若能使雄性虹鳟转化为雌性进行全雌养殖,到第3年再上市,则可延长有效生长期,达到大幅度增产的目的。牙鲆也有类似现象。
4.提高产品质量
大多数观赏鱼类中,雄性观赏鱼类比雌性具有更加艳丽的体色和外观,因此具有更高的商品价值。
5.提高性产品的产量
人们喜食带有蟹黄的雌蟹,造成膏蟹(雌蟹)和肉蟹(雄蟹)价格相差较大;
生产鱼子酱的鲟鳇鱼、卵价值较高的部分鲑科鱼类等,养殖全雌性群体可以成倍地提高养殖效率。
二、原始生殖细胞起源、迁移和分化(理解)
1.原始生殖细胞的起源
原始生殖细胞(来自内胚层生殖上皮)→ 生殖嵴(来自中胚层 )→ 生殖腺。
原始生殖细胞在进入生殖嵴之前,既可分化为精原细胞,又可分化为卵原细胞,这种分化是由其和不同的生殖嵴细胞的结合所决定的。这是性别诱导的基础!
2.原始生殖细胞的迁移
脊椎动物原始生殖细胞的迁移方式有3种:
①主动迁移:自身变形运动;
②被动迁移:通过周围组织细胞的分化生长运动推动;通过血液循环运动;
③化学动力迁移:即在将来形成生殖腺部位组织的诱导下, 被吸引过去。
3.生殖嵴的发生和分化
两栖类和其他高等脊椎动物:
生殖嵴原基=皮质(腹膜壁衍生)+ 髓质(中肾芽基衍生)。
①若皮质部发育,则髓质部退化 → 卵巢发生;
②若髓质部发育,则皮质部退化 → 精巢发生。
硬骨鱼类:生殖嵴仅由单一的胚性细胞组成,来源于腹膜壁。生殖嵴基质细胞与原始生殖细胞的相互作用,对鱼类性腺的分化至关重要。
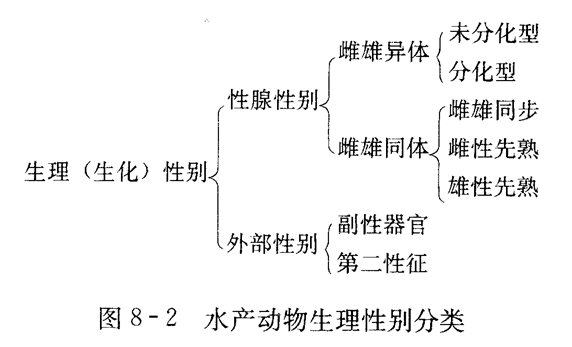
三、动物性别分类(理解)
1.生理性别:
由动物生理构成所表现出的性别,是动物性别的外在表现,是遗传性别与环境互相作用的结果;包括性腺性别和外部性别。
2.遗传性别:
由动物遗传基础决定的性别。有些鱼类及水产动物没有遗传性别,其性别的发育完全由环境所控制。
四、生理性别(掌握)
1.性腺性别:
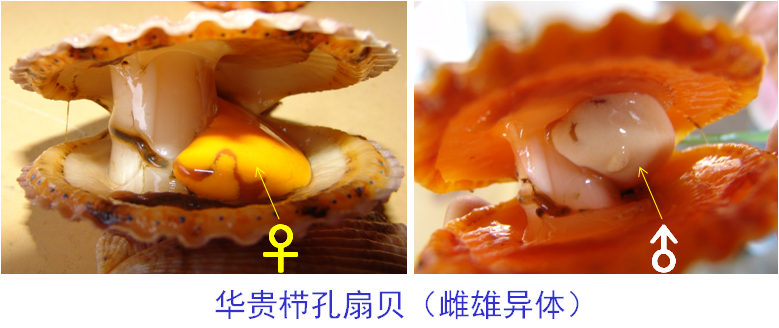
①雌雄异体:指一个个体内仅含有精巢或者卵巢,绝大多数鱼类属于这种性别。它还可以分为两类:分化型和未分化型。
A.未分化型:大多数鱼类,性腺首先发育成卵巢样的性腺,然后,大约一半的个体发育成雄性,另一半发育成雌性。
B.分化型:性腺直接分化成精巢或卵巢,如华贵栉孔扇贝。
②雌雄同体:雌雄同体是指同一个体中同时或依次存在着可以辨认的卵巢和精巢组织,分别产生卵子和精子。虽然可以产生雌雄性两种配子,但生物体通过一些机制限制自体受精的发生,多数情况下是卵子和精子的不同步成熟或不同步排放来避免。目前发现的雌雄同体鱼类大约有400种。
根据其雌雄功能出现的先后顺序可将其分为3类:
A.雌性先熟雌雄同体:由雌性功能转变为雄性功能的雌性先熟雌雄同体类型。低龄个体是一种性别(雌性),高龄个体则是另一种性别(雄性),例如石斑鱼类以及黄鳝均属这种类型。
B.雄性先熟雌雄同体:由雄性功能转变为雌性功能的为雄性先熟雌雄同体类型。早期卵巢的发育受到抑制,而精巢的发育较快,低龄鱼表现为雄性,只能排精不能产卵。随着年龄增大,精巢逐渐萎缩,卵巢逐渐发育成熟,表现为雌鱼。黑鲷、黄鳍鲷、金头鲷属于这一类型。
C.同步性成熟雌雄同体:雌雄两性同时存在,并同时具有雌雄两性功能,同时达到性成熟的雌雄同体称为同步性成熟雌雄同体类型。雌雄两性的性产物经不同的输出管道排出体外,一般情况下可以避免自体受精的发生。但有些种类存在自体的精子和卵子结合的现象。这种雌雄同步成熟的雌雄同体动物较少,多存在于较低等的水产动物之中,如墨西哥湾扇贝。

2.外部性别:
大多数水产动物的性别从外形上是很难分辨的,只有通过解剖和观察性腺才能确定。有一些种类在外部特征上有雌雄差异,这就是所谓的外部性别(external sex),或雌雄异形现象。
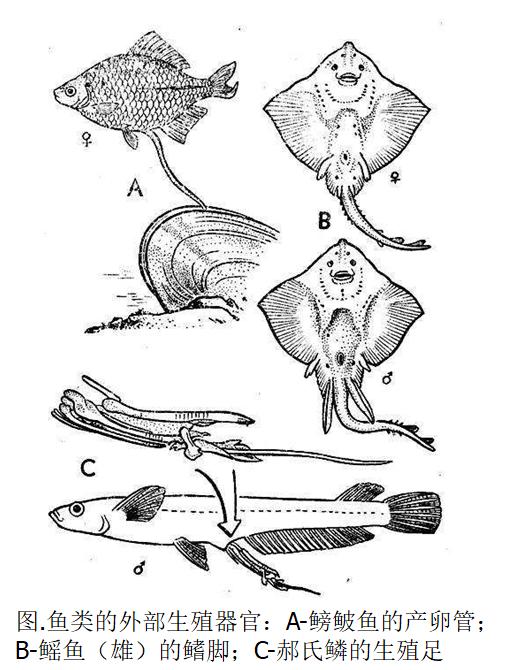
第一性征是指那些直接与繁殖活动本身有关的性征,即性附属器官。
例如,方斑东风螺的-交接器(下图);软骨鱼类雄鱼的-鳍脚,鳟科鱼类雄鱼的-生殖足,鲅虎鱼类雄鱼的-臀凸和鲚鲅类雌鱼的-产卵管等。
第二性征则是那些与繁殖活动本身无直接关系的性征,如雄鱼在生殖季节出现的婚姻装以及珠星等。

五、遗传性别(理解)
(一)鱼类遗传性别决定的特点
1.鱼类常染色体参与性别决定:
决定性别的基因并不明显地集中于性染色体上,常染色体上的基因也参与到性别决定中。通常性染色体是上位性的,但有时常染色体基因的作用可能超过它们,而且在不同种类,染色体在性别决定中的相对作用强度是不同的。证据包括:
①不同的雌性(雄性)与同一雄性(雌性)交配,得到不同性比的后裔;
②种间杂交得到不同的性比;
③环境因子对鱼类性比的影响;
④分子生物学证据。
2.环境因子影响鱼类遗传性别分化:
在发育早期,某些外部环境因素能在不同程度上影响鱼类的性别分化,从而使鱼类性别决定机制显得更加复杂。
3.部分鱼类性别由性染色体决定:
染色体性别(chromosomal sex):指遗传性别由来自卵细胞以及来自精子的染色体相互结合而决定的性别,相关染色体称为性染色体。
到目前为止,在2000多种真骨鱼中,被证明具有性染色体的种类很少,只有80种左右。其中能从细胞学上鉴别出性染色体的仅30余种,这说明绝大多数的鱼类性染色体异形的分化(下图)及性染色体尚处于低等的阶段。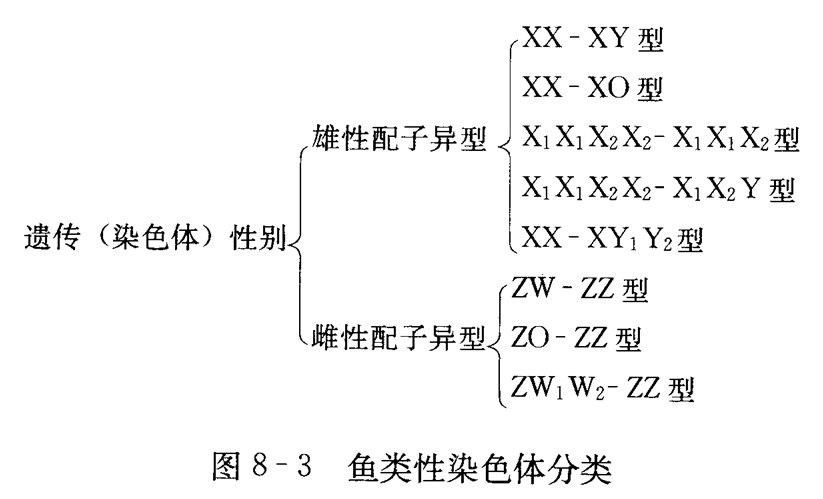
(二)鱼类性染色体的主要决定类型:
1.XX-XY型:
雄性是配子异型,可以产生两种不同类型的配子。性别由性染色体决定的鱼类大多数属于这种类型,如鲫、尼罗罗非鱼、胡子鲇和虹鳟等。多数高等哺乳类的性染色体也属此类。
2.XX-XO型:
雄性是配子异型,可产生两种类型的配子,但其中一种类型的配子缺少性染色体,用“O” 来表示。“褶胸鱼”即属这种性别决定方式,雌鱼的染色体有36条,表示为2A+XX=36,而雄鱼只有35条,表示为2A+XO=35。雄性产生的配子分别为:
A+X(n=18)和A+O(n=17)型。
3.ZW-ZZ型:
雌性是配子异型,可以产生两种不同类型的配子。大鳞副泥鳅、日本鳗鲡、奥利亚罗非鱼及食蚊鱼等均属这种性别决定方式。这也是鸟类常见的性染色体形式。
4.ZO-ZZ型:
雌性配子异型,在这种性别类型中,雌性有半数配子缺少性染色体。属这种性染色体类型的鱼类较少。洪云汉(1984)报道短领鲚为此类型。
六、鱼类的性转变(理解)
1.鱼类的性转变现象
性逆转:是雌雄异体的动物由一种性别向另一种性别转变的过程,其结果是动物体性别的完全转变,过程可以具有雌雄两种性腺或性腺退化到未分化期,然后性腺再重新分化。
性逆转现象有两种可能:一是生理性的;二是病理性的。
鲷科鱼类中的平鲷、黑鲷、黄鳍鲷、金头鲷等在低龄是雄性,高龄转变为雌性,属雌雄同体雄性先熟种类;酯科鱼类中的石斑鱼则相反,低龄时是雌性,高龄转变为雄性,属雌雄同体雌性先熟种类。
2.影响鱼类性转变的因素
①激素与性转变
脊椎动物遗传性别的形成先于生理性别,从遗传性别到生理性别的形成是一个复杂的过程。
在这个过程中,遗传基础起了重要的作用,控制性别发育的方向,然而当环境或其他因素产生变化,就有可能通过激素变化影响个体性别发育的方向。
雄激素能够诱导雄性化,雌激素可以诱导雌性化。生理性的性反转可能是通过基因对激素的产生进行调控而完成。
②环境与性转变
尼罗罗非鱼和奥丽亚罗非鱼在高温(34-37℃ )时,产生较高的雄性率,而莫桑比克罗非鱼则在低温(19或20℃)时能增加雄鱼的比例。
丝鳍花酯“社会控制”:一般20尾左右一群,每群有一尾雄鱼,其余都是雌的。当移走雄鱼后,则剩余的最强健的一尾雌鱼会发生性转变,成为雄鱼。如此继续移走雄鱼,则最后一尾雌鱼都会变成雄鱼。
3.鱼类性转变机制
①去抑制机制:认为,在繁殖过程中伴随卵子发育而分泌的前列腺素刺激下丘脑,促进垂体释放促黄体激素(LH)和促卵泡激素(FSH),从而诱导卵巢提前转变成精巢。
②刺激机制:认为,当雄鱼与雌鱼相互间的特异联系方式改变时,雌鱼的高级神经中枢就会将这种改变的信息转换成下丘脑的刺激因素,下丘脑再促使垂体释放性转变因子。
这两种机制的共同特点是它们都涉及中枢神经系统“下丘脑-垂体-性腺轴”。因而,神经内分泌途径可能是外部刺激与性转变的内在变化之间的一座桥梁。
复习题:
一、问答题:
1.试述原始生殖细胞的起源、迁移和分化。
二、简答题:
1.水产动物性别控制有什么意义?
2.水产动物性别是如何分类的?
3.水产动物生理性别是如何分类的?
4.水产动物性腺性别如何分类?
5.水产动物外部性别如何分类?各有什么特征?
6.鱼类遗传性别决定的特点是什么?
7.鱼类性染色体如何分类?
8.鱼类性染色体的主要决定类型有哪些?
9.举例说明影响鱼类性转变的因素。
10.鱼类性转变的机制是什么?
三、名词解释:
1.生理性别;
2.遗传性别;
3.外部性别;
4.染色体性别;
5.性染色体;
6.性逆转;

