

第七章 雌核发育与雄核发育
第24课时 雌核发育
一、雌核发育现象(理解)
1.雌核发育(gynogenesis):
是指受精后,卵子在精子的刺激下,依靠自身的细胞核发育成个体的一种有性生殖方式。精子仅起激活作用,不与雌性原核融合,所产生的后代完全与母本相似。
2.非减数卵子(non-reductional egg):
雌核发育种类产生 的卵子,其染色体数目仍与体细胞的染色体数目相同,称之。
3.天然雌核发育:
排出的卵子处于第一次成熟分裂中期,与 异源精子受精,放出第一极体,精子激发卵子进行雌核发育,未发生核融合,卵子不进行第二次成熟分裂,从而获得具有与母本完全相同的染色体组型。
雌核发育的鱼类多为三倍体。我国对鱼类天然雌核发育研究得比较清楚的是黑龙江水系的银鲫。大量的研究表明,其精卵形成机制是相当复杂的,基本上通过不发生第一次成熟分裂、成熟分裂后第二极体不排出、第一次卵裂为核内有丝分裂等来维持其倍性。
4.人工雌核发育(artificial gynogenesis):
是指用遗传失活的精子激活卵子的发育,精子不参与合子核的形成,卵子仅靠雌核发育成胚胎的现象。
人工雌核发育的胚胎是单倍体,孵化时呈现显著的畸形,没有存活能力。通过加倍卵子的染色体使其恢复二倍性后,便成为外观正常的具有存活能力的雌核发育二倍体(gynogenesis diploid)。
二、精子染色体的遗传失活方法(理解)
1.γ射线和X射线
具有较好的穿透能力,可诱发染色体断裂。当照射达到一定剂量后,能够使精核中的DNA完全被破坏。
Hertwig效应:
采用物理射线处理精子时,随着照射剂量的增高,受精后胚胎的成活率降低;当照射超过某一剂量时,成活率反而回升,这种现象称作Hertwig效应(Hertwig effect)。
解释:照射后的突然变异和染色体切断对胚胎产生毒害作用;仅靠由卵子来的染色体发育时,成活率回升。
作用:通过观察Hertwig效应可以推定射线照射精子的适宜剂量。
2.紫外线(UV)
紫外线是一种相对安全的射线。精子DNA经紫外线照射后形成胸腺嘧啶二聚体,使DNA双螺旋两链间的氢键减弱,从而使DNA结构局部变形,阻碍DNA的正常复制和转录。
在诱导斑马鱼、狗鱼、鲈、鲇和鲤雌核发育的过程中,均使用过紫外线照射作为失活精子的手段。
美洲牡蛎、太平洋牡蛎、皱纹盘鲍、栉孔扇贝、虾夷扇贝等全部使用紫外线照射作为失活精子染色体的手段。
3.化学试剂
采用甲苯胺蓝、乙烯脲、二甲基硫酸盐 、吖啶黄和噻嗪等化学药物诱变精子DNA的方法也已经用于诱导鱼类与两栖类雌核发育的研究中。最适处理浓度和处理时间还需进一步进行研究。
4.杂交诱导
鲽×拟庸鲽、川鲽×拟庸鲽、银大麻哈鱼×溪红点鲑、草鱼×鲤→放出两个极体后→雌核发育单倍体胚胎。
形成单倍体原因:胚胎细胞分裂节奏不同步;不亲和性导致雌原核对雄原核的排斥。
吴仲庆(1983)在研究两栖类种间杂交诱发雌核发育的机制时认为,异质精子穿人卵内后,精核不参与发育,但是近侧中心粒向卵子提供了精子星体,从而提供了雌核发育所必需的双星光,使个体发育得以进行。
中心粒:是中心体的主要结构,位于核附近由9组三联体微管围成的成对圆筒状结构,即一个中心体含有一对中心粒,且互相垂直形成"L"形排列。

有丝分裂器:由动物细胞的中心粒、星体、纺锤体、染色体等组成了。动物细胞借助有丝分裂器的作用,使染色体能够准确地,不有条紊地在细胞内活动,从而使细胞正常地进行分裂。有丝分裂器有两极,每极有一对中心粒。分裂器的极,决定了染色体的运动方向;分裂器赤道面的方向,决定了母细胞分裂成两个子细胞横缢面的位置。有丝分裂器还能把成对的染色体拉向相反的两极。
三、雌核二倍体的人工诱导
1.诱导原理图(掌握)
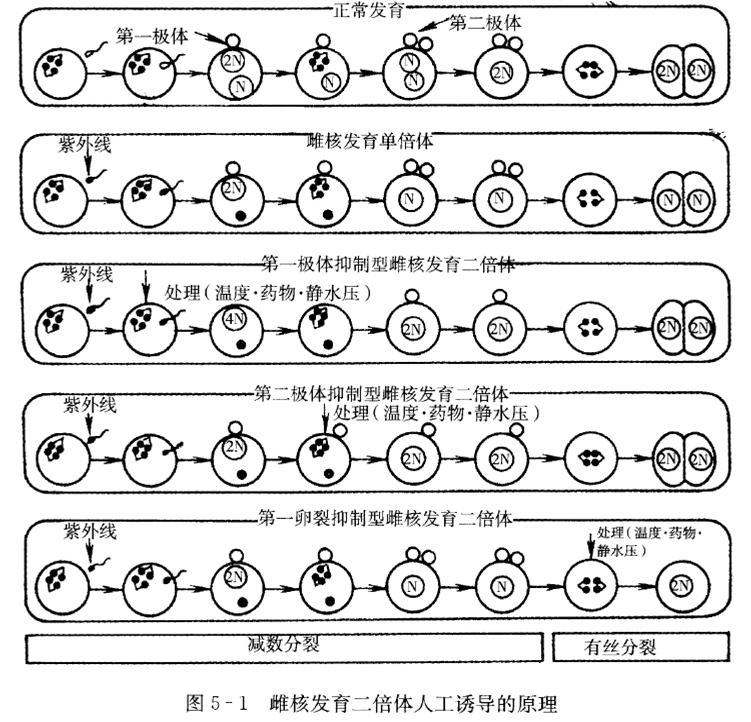
2.雌核发育二倍体研究情况(了解)
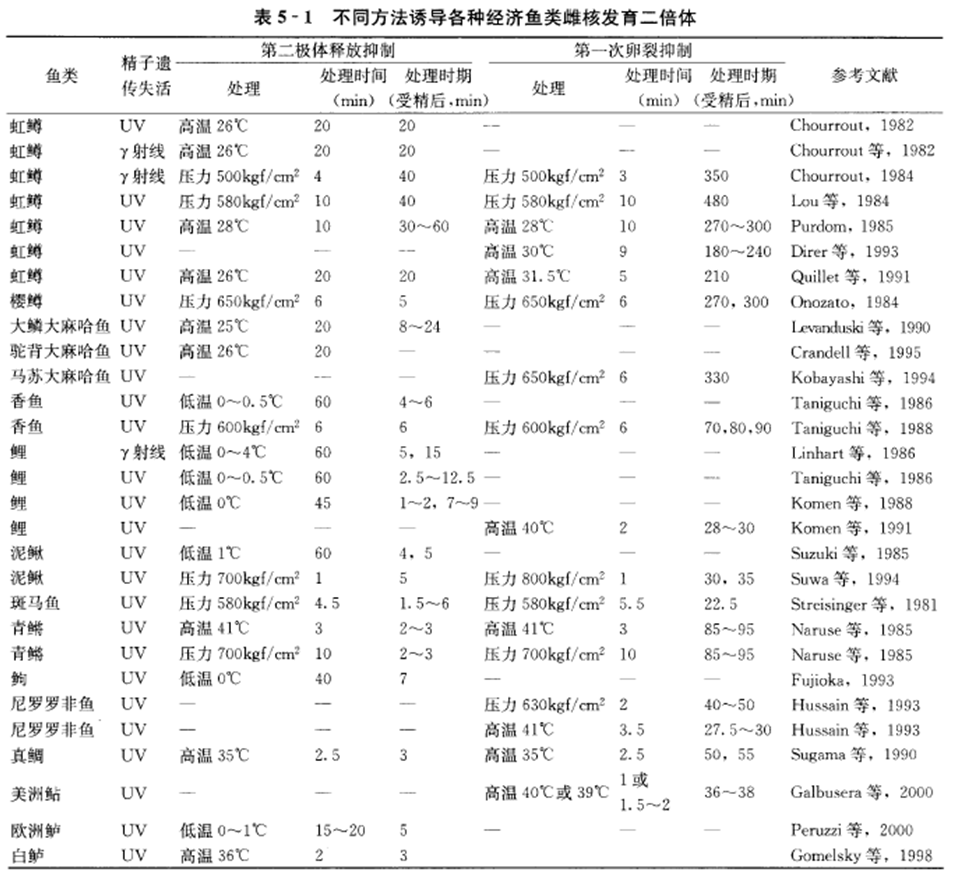
①迄今已获得人工雌核发育二倍体的鱼类有30多种(表5-1)。
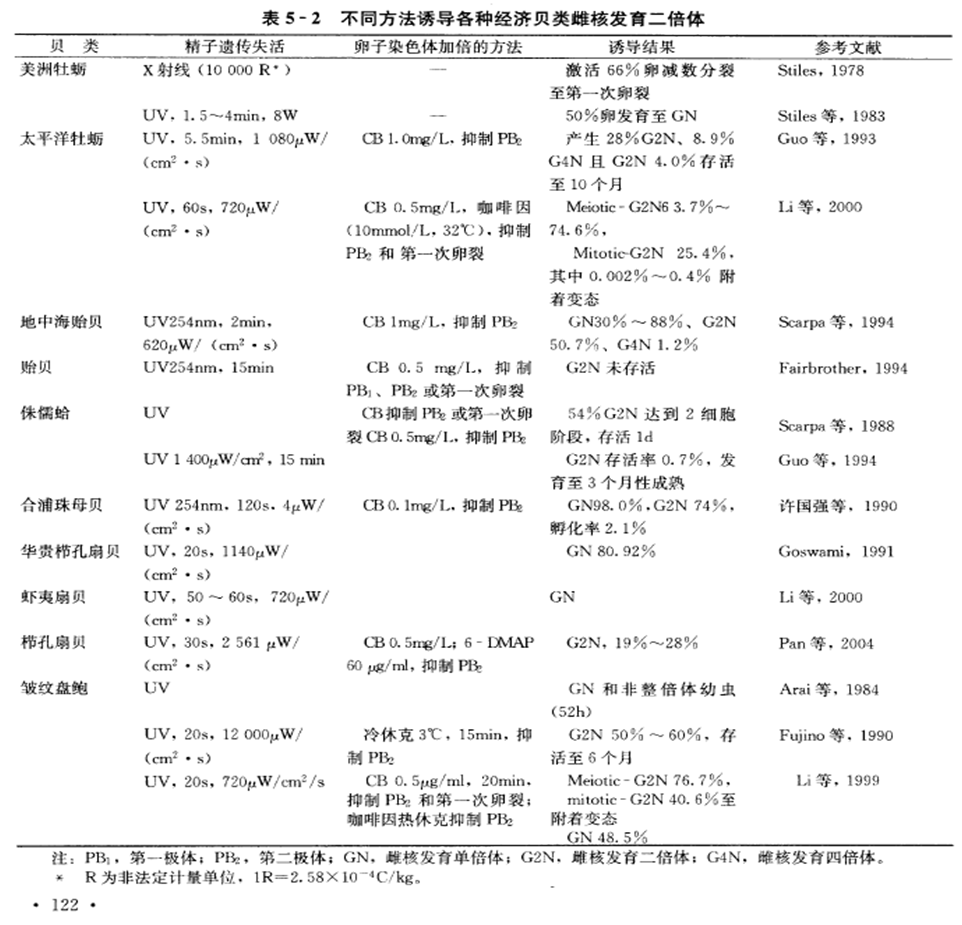
②在海产经济贝类方面,人工雌核发育的研究开展较晚,至今还没有培育到成体的报道。据统计,目前已在10种海洋贝类进行了人工诱导雌核发育的研究(表5-2),其中双壳类9种,腹足类1种。
③甲壳类雌核发育诱导目前仅在中国对虾有报道。
3.雌核发育二倍体的缺点(理解)
①存在大量非整倍体:父本染色体部分失活,导致父本染色体不同程度地参与核分裂,从而产生非整倍体。
②成活率明显降低:卵子染色体加倍造成隐性致死或有害基因的纯合,使诱导出的雌核发育后代难存活,也有可能是由于精子照射造成的遗传损伤引起存活率下降。
四、雌核发育二倍体的鉴别(理解)
当雌核发育二倍体产生的时候,必须有充分的证据证明染色体数目的二倍性,且精子DNA确实没有参与胚胎发育。
1.流式细胞仪检测法
这种方法具有快速、准确地分析大量样品,且取样量少等优点,但也有一定的缺陷:流式细胞术不能提供所测个体染色体组成的资料,且在技术上较难检测出DNA小剂量的差别,其显示的整倍体个体可能包括增加或减少了几条染色体的非整倍体。
2.荧光显微分析法
与流式细胞仪检测法相似,所需仪器比流式细胞仪便宜,但需要借助测光设备对细胞逐个地进行测量,无法像流式细胞仪检测法那样短时间内测量大量细胞。
3.染色体分析法
染色体的计数和组型分析对鉴别雌核发育是最为直接和准确的方法。
五、雌核发育二倍体受精细胞学机制(理解)
1.精核受到抑制:
在鱼类的人工诱导雌核发育研究中,一般认为入卵后的精核始终受到抑制而不形成雄性原核,保持固缩状态,不与雌性原核融合,精核仅起激活卵子启动发育的作用。
2.核膜解体异常:
关于精核在银鲫卵质中不能形成雄性原核的原因,楼允东(1999)认为这是由于在雌核发育受精状态下,银鲫卵质促进精核核膜解体的功能异常,使覆盖精子头部的核膜不能像在两性融合生殖受精卵子中那样进行崩解,所以精核不能进一步发育为雄性原核。
3.形成染色质小体:
Li等(2000)在对太平洋牡蛎和皱纹盘鲍的雌核发育二倍体的研究中发现在第一卵裂中期,雌核发育卵子内的雄性原核并不能浓缩形成染色体,而是形成致密的染色质小体(dense chromatin body,DCB),位于母本染色体近旁,没有与母本染色体融合。在第一卵裂后期,或随着卵核由来的一组染色体移动,或滞留在2组卵核由来的染色体之间伸长。当胞质分裂结束后,可以观察到DCB存在于2个分裂球其中之—的细胞质中,或被第一卵裂沟分割为2个部分而同时存在于2个分裂球的细胞质之中。
六、雌核发育二倍体的生物学特性(理解)
1.性别
① 全雌性(XX型):自然界大部分鱼类是雄性异型(XY型,XX(♀)— XY(♂))种类,雌核发育后代全雌性(XX型)。 Guo等(1994)在进行侏儒蛤的雌核发育诱导时发现,侏儒蛤中雌性是同型配子(XX),性别遗传机制为XX(♀)— XY(♂)。
②雄性(ZZ)和雌性(WW)各占一半:对于雌性异型(ZW型,ZW(♀)-ZZ(♂))种类,雌核发育后代雌雄各占一半。因此,通过分析雌核发育后代的性别比例就可以了解该动物的性别决定机制。Avtalion等(1990)发现奥利亚罗非鱼雌核发育获得的后代有雌有雄,从而得知该鱼的性别遗传机制为ZW(♀)-ZZ(♂) 。
2.繁殖力
吴清江等(1999)认为通过物理或化学方法加倍的鱼类雌核发育个体,大约有80%的性腺不能正常发育,而染色体不完整是人工雌核发育鱼类不育的主要原因。
邓岳松(1999)观察了人工雌核发育草鱼性腺的早期发育情况,所有个体都是雌鱼,大部分性腺发育正常,少数性腺发育畸形或迟缓。
根据已报道的研究结果,雌核发育二倍体侏儒蛤性腺发育至3个月后大部分个体均可观察到配子,并且其怀卵量(2.4×105)比普通二倍体(3×105)的少,相对生殖力为79%,卵径(48.6± 3.1um)与普通二倍体的卵径(49.1±3.1um)相当。
3.生长与发育
总的来说,雌核发育所产生的后代与正常受精所得到的胚胎相比发育速度变慢,成活率低。Scarpa等(1994)报道地中海贻贝雌核发育二倍体幼虫的个体大小总是小于对照组,且其附着变态的时间也迟于对照组大约3周。
Fujino等(1990)报道皱纹盘鲍雌核发育二倍体与正常二倍体相比,在大小上存在较大差异。
Guo等(1993)报道太平洋牡蛎雌核发育二倍体发育至8月龄时,与对照组之间在大小上无太大差别。
七、雌核发育二倍体的遗传学特性(理解)
杂合型亲本的第一极体抑制型(如图A)和第二极体抑制型(如图B):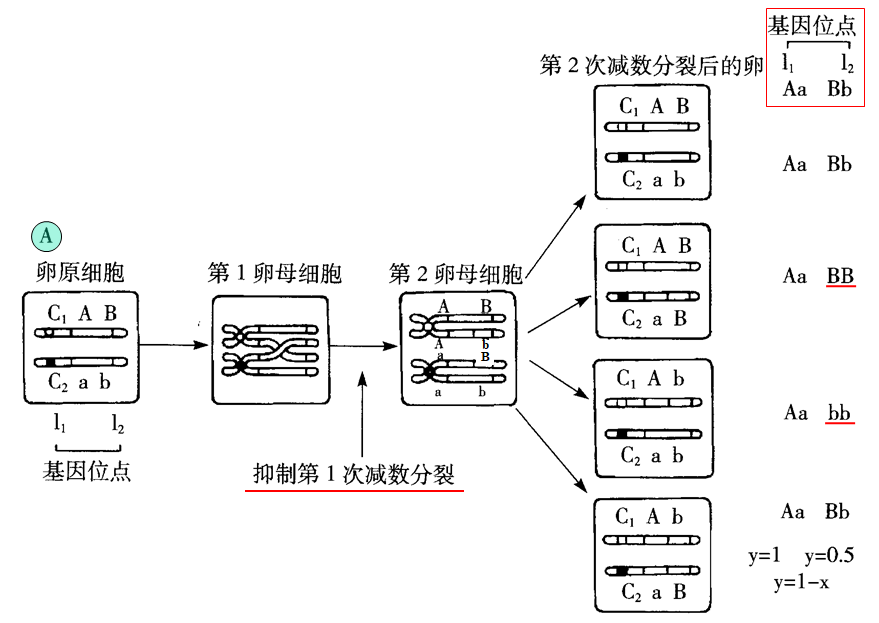
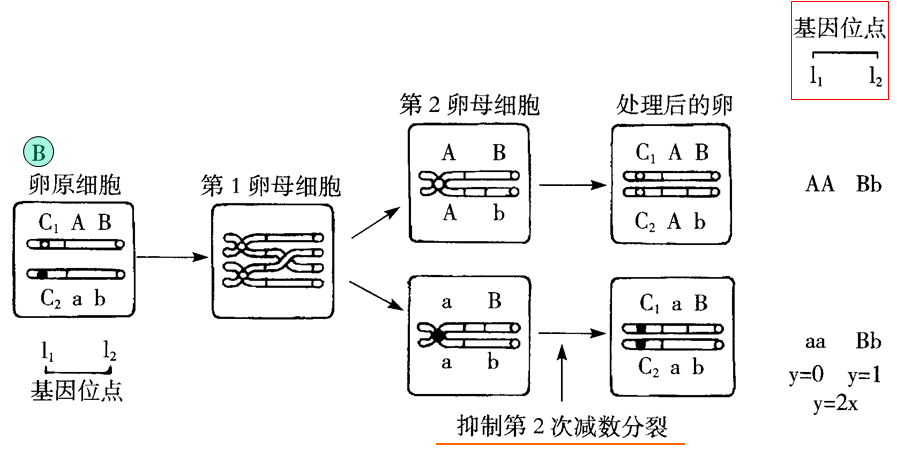

八、雌核发育二倍体在育种中的应用(理解)
1.培育特殊品种
异精雌核发育(allogynogenesis):在用兴国红鲤为父本、方正银鲫为母本的杂交实验中,发现异源精子不仅能刺激银鲫雌核发育,而且还能影响子代的生长、性比、体色等性状,故而把这种表现出异源精子生物学效应的雌核发育称之为异精雌核发育。银鲫异精雌核发育的子代统称为异育银鲫。异育银鲫具有明显的生长优势,已在全国许多地方推广并取得了较高的经济效益。
异精效应的解释:异源精核可能与银鲫之间发生了极为有限的遗传物质交流,发生DNA杂交。在分子遗传学上也找到一些相关的证据。
2.快速建立纯系
在人工雌核发育中,阻止第一次卵裂产生的雌核发育二倍体,由于所有的基因都处于纯合状态,成为真正的纯系,下一代如重复雌核发育即可以产生克隆种群。
克隆种群的形成,使过去需要几十年、几百年才能完成的纯系培养,甚至在理论上需要无数次兄妹交配才能完成的完全纯系培育在几年内就可实现,所以在水产动物育种中有着极高的应用价值。
3.性别控制和单性种群利用
雌性鲤:生长速度要比雄性的快15%-25%,因此,通过培育全雌鲤可以提高产量;
雌性对虾:可以大大提高对虾养殖的产量;
雌性洄游性鲟:鱼籽(雌性生殖腺)有极高的经济价值,养殖全雌鲟的经济效益比雌雄混合群体养殖的要大得多。
全雌草鱼:Stanley等(1975)通过人工雌核发育的方法培育,达到既能清除河道大量水草又能控制其过量繁殖的目的。
4.确定性别遗传机制
由于水产动物的染色体数量多,个体小,雌雄个体之间难以找到具有形态差异的染色体,更难以确定性染色体,因此要确定性别决定机制是属于雌性同配还是雌性异配颇为困难。采用雌核发育的办法,只需确定雌核发育子代的性别即可得知。
5.基因一着丝点作图
在人工诱导的雌核发育中,通过计算杂合后代在群体中的比例,就可以推算基因重组频率,根据基因重组频率可以确定基因在染色体上的位置及基因间的距离,从而能绘制基因连锁图。
复习题:
一、问答题:
1.试述雌核发育二倍体的生物学特性。
2.画图说明杂合型亲本第一极体抑制型雌核发育二倍体遗传学特性。
3.画图说明杂合型亲本第二极体抑制型雌核发育二倍体遗传学特性。
4.雌核发育二倍体在遗传育种中有什么应用?
二、简答题:
1.什么叫Hertwig效应?为什么有这种现象?有什么作用?
2.在人工雌核发育中,让精子染色体遗传失活的方法有哪些?
3.画出人工诱导雌核发育单倍体原理图。
4.画出第一极体抑制型人工雌核发育二倍体原理图。
5.画出第二极体抑制型人工雌核发育二倍体原理图。
6.画出第一次卵裂抑制型人工雌核发育二倍体原理图。
7.简述雌核发育二倍体研究情况。
8.雌核发育二倍体有什么缺点?
9.雌核发育二倍体的鉴别方法有哪些?
10.介绍雌核发育二倍体的受精细胞学机制。
11.什么叫异精雌核发育?如何解释这一现象?
三、名词解释:
1.雌核发育;
2.非减数卵子;
3.天然雌核发育;
4.人工雌核发育;
5.异精雌核发育;
6.异育银鲫。

