

第23课时 多倍体的应用
一、多倍体的鉴定(掌握)
1.染色体计数法(贝类染色体制片法)
①用担轮幼虫制备染色体标本;
②用鳃组织制备染色体标本;
③用上足触手、外套膜制备染色体标本。
.png")

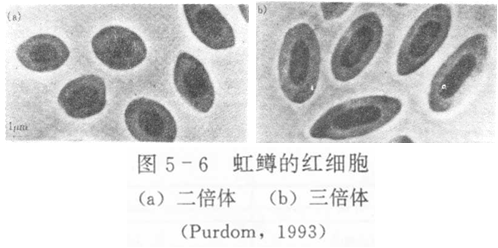
2.细胞核体积测量
一般认为,真核细胞的核大小与染色体数目成正比,同时细胞核与细胞质在细胞中总是维持较稳定的核质比。
核面积=πab/4;
核面积比: 二倍体︰三倍体=1:1.3
核体积=(4/3)πab2;
红细胞核体积比:二倍体︰三倍体=1:1.5;
二倍体︰四倍体=1:1.74;
式中,a为短轴长度,b为长轴长度。
目前,红细胞的测量被广泛用来鉴定多倍体鱼。不过,也有学者认为这个方法不能准确地反映染色体倍性。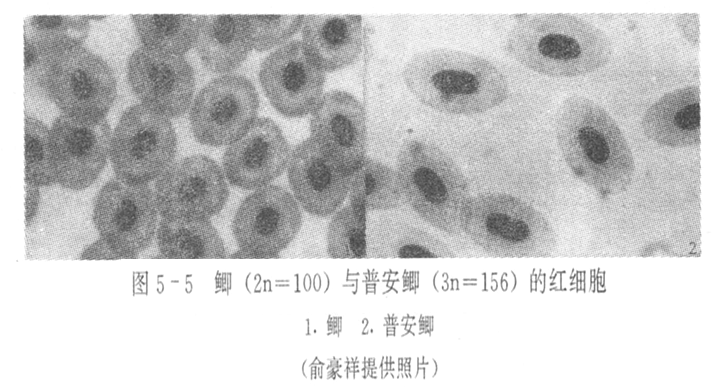
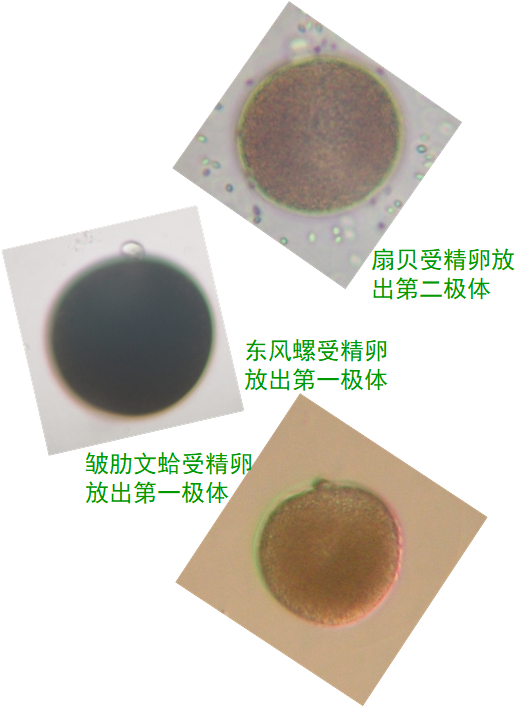
3.极体计数法
因为三倍体的诱导是通过阻止第一极体或第二极体的排出而完成的,所以三倍体合子只有一个而不是两个极体。
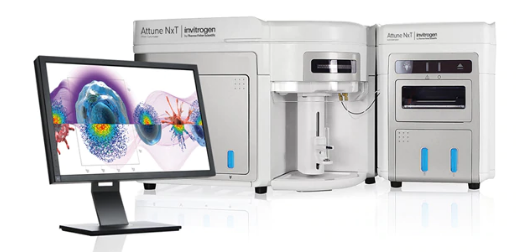
4.DNA含量测定方法
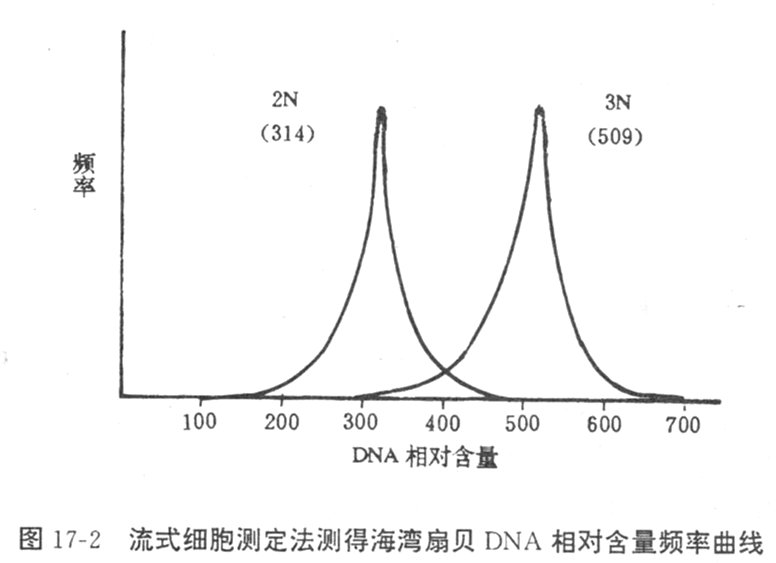
①流式细胞仪检测
用DNA-RNA特异性荧光染料(如DAPI)对细胞进行染色,在流式细胞仪上用激光或紫外光激发结合在DNA上的荧光染料,检测每个细胞的荧光强度,其强度与DNA含量成正比,与已知的二倍体细胞或单倍体细胞(如同种的精子)荧光强度对比,判断被检查细胞群体的倍性组成。
广泛应用于鱼类、对虾和贝类等。方法:抽取鲍鳃基血窦的血液0.02~0.04ml,注射到预先装有0.05~0.1ml超滤海水的测试小管中,加入1.0~1.5ml DAPI染液,上机进行测试。
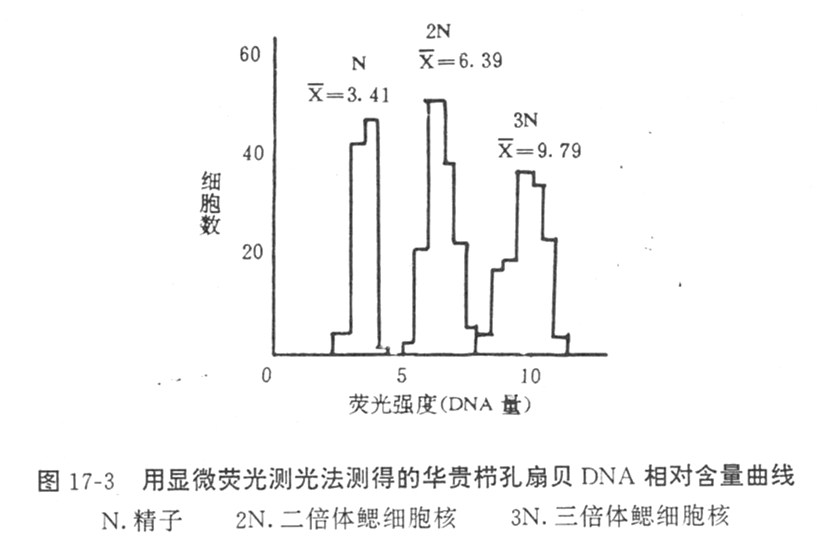
②显微荧光测定法
将被测材料用固定液固定后,将组织用细胞匀浆仪弄碎,使细胞游离,制成涂片标本,或者采取血液制成血涂片。标本用DAPI染色,然后在显微分光光度计上,通过波长365nm的紫外线照射,用扫描测光装置测定细胞核的荧光强度,获得处理组个体和对照组个体之间细胞核DNA相对含量曲线,从而测出二者的DNA比率,即可确定处理组个体的倍性,进而计算出三倍体的诱发率。Komarv(1987)采用这种方法测出华贵栉孔扇贝的三倍体诱导率。

二、多倍体的生物学特性(掌握)
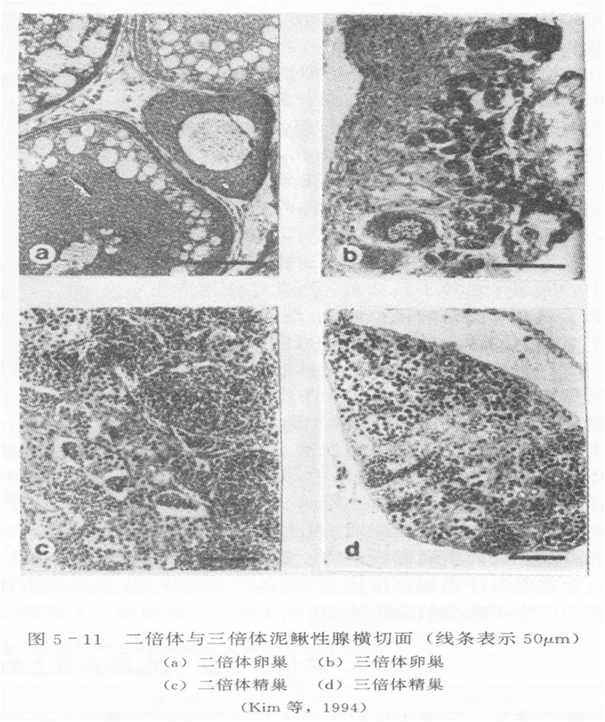
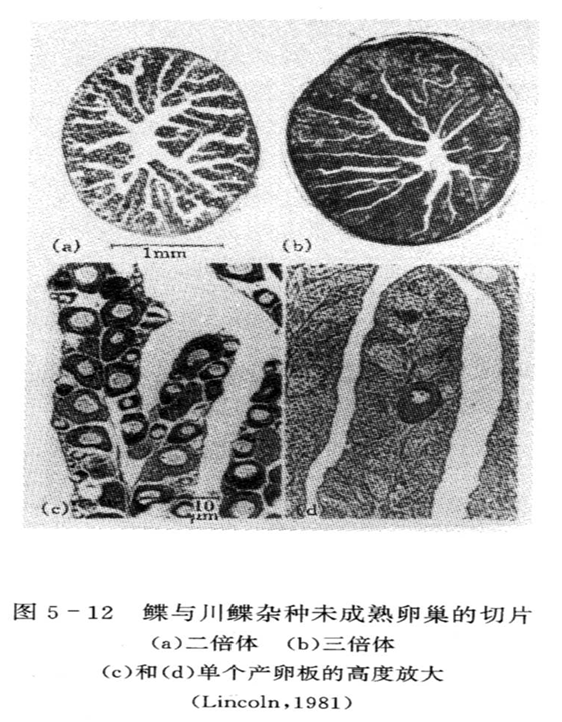
1.性腺发育
人工诱导三倍体贝类的性腺发育大致有3种类型:
①大部分不能发育:海湾扇贝、砂海螂、华贵栉孔扇贝。
②有不同程度发育:长牡蛎、美洲牡蛎能形成精子和成熟的卵母细胞;杂色鲍能产生成熟的卵母细胞,但不能形成成熟的精子。
③产生的精、卵能与二倍体交配或三倍体自交,但受精卵发育到D形幼虫就大量死亡,如三倍体珠母贝。
此外,四倍体个体的性腺发育正常,但育性较二倍体表现差。
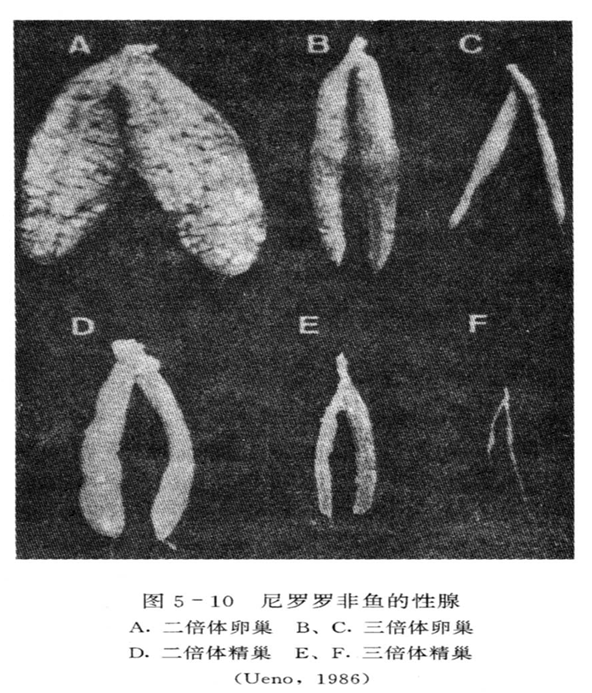
2.生活力与生长
①生活力:大多数三倍体鱼类具有正常的生活力,但虹鳟和银大麻哈鱼等人工诱导三倍体的生活力比二倍体差一些。
②生长:在早期生长上,有的三倍体鱼要比二倍体快;更多的研究表明三倍体和二倍体两者在早期生长方面无明显差异。多数研究认为,贝类达到成熟期后,三倍体才显示出生长优势,表明三倍体用于性腺发育的能量转而用于体细胞的生长。
3.其它性状
①三倍体虹鳟的肉质优于二倍体;
②三倍体大西洋鲑耐低氧能力高于二倍体,故可适于低氧环境养殖;
③有的情况下三倍体的个体不比二倍体大,这必然伴随三倍体个体细胞数量的减少,同时也伴随各个器官细胞数量的减少。
三、多倍体的用途(理解)
1.提高生长速度
①孙振兴等(1992)发现皱纹盘鲍三倍体与二倍体的当年稚鲍在壳长和体重增长上无明显差别;2龄后期,三倍体的壳长和体重生长才逐渐优于二倍体;3龄时,三倍体的体重、软体部和足肌的增长都明显快于二倍体。
②美国的海湾扇贝三倍体经114天(16周)的试养后,闭壳肌重量平均比二倍体高73%,软体部湿重平均比二倍体高36%,闭壳肌指数和肌肉糖元含量也比二倍体高(Tabarini,1984)。
③华贵栉孔扇贝三倍体经13个月的养殖,肉重和闭壳肌重分别比对照组的二倍体多39.5%和67.4%(林岳光等,1995)。
④日本对三倍体牙鲆的开发已进入实用阶段,普通牙鲆上市需经3年,而三倍牙鲆上市只需2年,三倍体的生长速度是二倍体的1.4倍,存活率也较高(钟正新,1988)。
⑤近江牡蛎三倍体比二倍体生长快、糖原含量高、成熟期后存活率高于二倍体。
2.控制繁殖
①利用三倍体进行引种试验、确定生态安全性后再引进2N;
②可以通过养殖不育群体控制养殖密度;
③用三倍体群体完成一些短期的目标,目标完成以后群体也自然消亡。如在杂草丛生的水体放养不育的草鱼三倍体,既能除草又可防止因其繁育后代导致的生态变化。
3.延长寿命
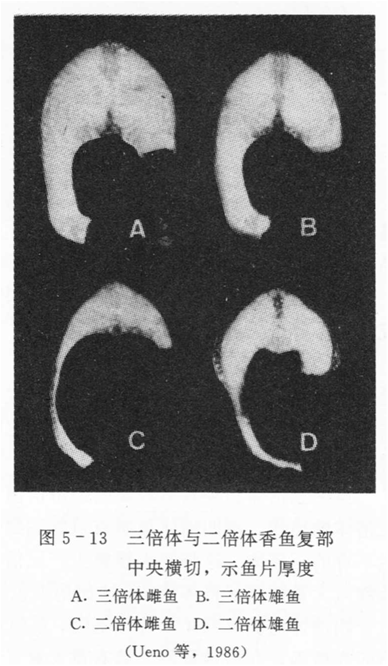
对于那些在性成熟时会遭受损失或有成熟死亡的鱼类来说,不育三倍体可能比二倍体存活得更长久。香鱼的寿命很短,一般是在秋季产卵繁殖,而后结束其一生,而三倍体香鱼可以跨年生长,还可以全年上市销售。
4.改善品质
①口感好:三倍体物种的细胞体积大,因此同样大小的二倍体和三倍体,三倍体的细胞数量要少于二倍体,相对的筋膜等也少于二倍体,并且三倍体的鱼肉看起来较有光泽,因此三倍体个体的肌肉的口感柔软,更受消费者的偏爱。
②肉质美:性成熟的鱼类往往伴随着肌肉质量的下降,所以欧美国家消费虹鳟等鱼类时,偏好个体较小的未成熟个体。三倍体的鱼类因性腺不发育,生长中没有肌肉质量下降的问题(如香鱼,下图)。
四、多倍体研究进展(掌握)
(一)美国牡蛎三倍体研究情况
1.概况:
70年代末开始,80年代中期应用于生产:
85年 3N 100万个
87年 3N 20~30亿
88年 3N 120亿(10万亩种苗)
占美西北沿岸牡蛎苗种产量的1/2。
研究品种:C.gigas 、C.virginica
推广地区:西北部沿岸发展至东海岸
美国渔业界认为:肉质好、个头大的3N牡蛎已进入夏季市场。这改变了夏季从亚洲高成本进口牡蛎的做法。
2.3N诱导实验
①方法:采用细胞松弛素C.B抑制受精卵第二极体放出,形成3N;
②药物浓度:1L+0.5~1g DMSO+0.5~1mgC.B;
③试验温度设置:18、20、25℃;
④处理密度:300~500、5000、1亿/L;
⑤受精后开始处理时间:0、15、30、120分钟;
⑥处理持续时间:15分钟,过程搅动;
⑦处理后清洗:25um网滤,浸入0.1%DMSO海水溶液15分钟,不停搅动,再移入原温度组中搅动,水体150L。
3.试验结果:
①25℃、30分钟开始处理,诱导率达88%;
②密度以5000万粒/L为好;
③CB浓度(mg/L):
0.5mg/L-77%( 5000万粒/L )
1.0mg/L-87%( 5000万粒/L )
0.5~1.0mg/L-30%(1亿粒/L )
4.3N的鉴定技术-荧光显微镜、流式细胞仪:
原理:细胞对荧光染料(DAPI)进行吸收,吸收量与DNA含量成正比。
优点:操作简便快捷,可活体取样(鳃丝),取样量少,效率高,在美国得以迅速推广。
5.3N牡蛎预期效果的印证:
①对人体无害;
②生殖季节肉质好;
③第2年出现生长优势。

(二)我国牡蛎三倍体研究情况
1.化学诱导获得三倍体
参考美国牡蛎三倍体研究方法,但诱导率难以稳定达到70%以上的水平,这与卵子同步成熟技术不过关有关,影响三倍体的产业化。
2.杂交法获得三倍体
目前国内流行趋势是用四倍体与二倍体杂交产生100%的三倍体,太平洋牡蛎及葡萄牙牡蛎三倍体规模化养殖已在山东、福建一带取得成功。四倍体的成功诱导和养成,并具有繁殖四倍体后代的能力,是获得100%三倍体的关键。2019年,葡萄牙牡蛎三倍体养殖区域从福建拓展至广东粤东至粤西沿海,甚至延伸至广西和越南。目前,要获得具有推广意义且占比达到100%的牡蛎三倍体,采用四倍体与二倍体杂交是唯一的途径。如何获得可以繁殖后代的四倍体是这一途径的关键点。
(1)四倍体的诱导:
①由二倍体直接诱导四倍体
关于贝类四倍体的诱导方法,已有报道用普通二倍体卵子授精,并通过抑制第一极体、第一和第二极体或第一次卵裂,以及采用细胞融合,人工雌核发育结合抑制两个极体等方法来诱导四倍体,但这些方法都没能获得可存活的四倍体成贝。
1994年Guo和Allen提出了长牡蛎四倍体胚胎采用上述方法不能存活的原因在于长牡蛎受精卵发育类型是镶嵌型发育,由二倍体卵子授精后,经人工诱导处理形成四倍体核的细胞,由于四倍体细胞核较大,细胞质不能满足四倍体核完成所需的分裂次数,造成分裂细胞数目不足(导致器官发育不齐全)而引起的。认为如果能增大卵子的体积,就可能解决这个问题。
镶嵌型发育:
如果在发育早期将一个特定分裂球从整体胚胎上分离下来,他就会形成如同其在整体胚胎中将会形成的结构一样的组织,而胚胎其余部分形成的组织会缺乏分离裂球所能产生的结构,两者恰好互补。这种以细胞自主特化为特点的胚胎发育模式称为镶嵌型发育。如:栉水母、海鞘、环节动物、线虫、软体动物。
调整型发育:
对细胞进行有条件特化的胚胎来说,如果在发育早期将一个分裂球从整体胚胎上分离下来,剩余胚胎中某些细胞可以改变发育命运,填补分离掉的裂球所留下的空缺,仍形成一个正常的胚胎。这种以细胞有条件特化为特点的胚胎发育模式称为调整型发育。
②由三倍体诱导四倍体
因此Guo和Allen采用了体积显著大于二倍体卵子的三倍体卵子(注:少量三倍体可以形成成熟的性腺)与正常二倍体精子授精后,用CB抑制第一极体排放,首次成功地培育出可存活的长牡蛎四倍体。而这种诱发四倍体的机制,提出的假设是第一极体受抑制的三倍体卵子进行第二次减数分裂时发生了联合两极分离,30条染色单体参与形成第二极体而排放到卵子外,另外30条染色单体留在卵子中与10条精子染色体结合,产生了四倍体。Que等(1997)观察了长牡蛎三倍体卵子受精后第一极体被抑制的染色体分离行为,从而在细胞学方面证实了这个假设。
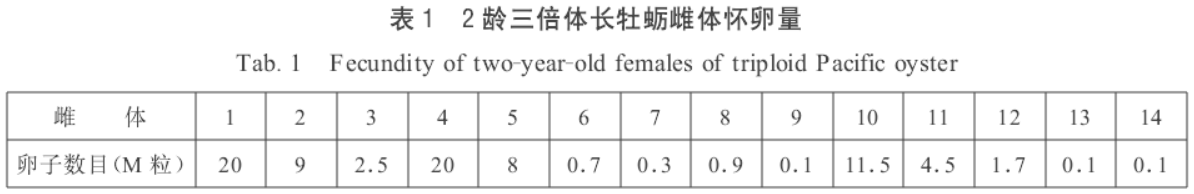
林琪等采用了这一技术方法,具体操作:
用剖取法取得精卵,卵子经200目筛绢滤去组织块,在500目筛绢框里经砂滤海水清洗后,授精,受精处理的水温26~27℃ ,海水比重1.021。三倍体个体怀卵量:
处理组用三倍体卵子与二倍体精子受精,并在受精后8min,用0.5mg/dm3 CB处理受精卵25min以抑制第一极体排放,在含0.1% DMSO(二甲基亚砜)的海水中浸洗2次,移至500dm3容器中孵化,收集上浮担轮幼虫进行培育至D型幼虫,培育密度2个/cm3,常规育苗。对照组用三倍体卵子与二倍体精子授精,但不经过CB处理。结果如下:
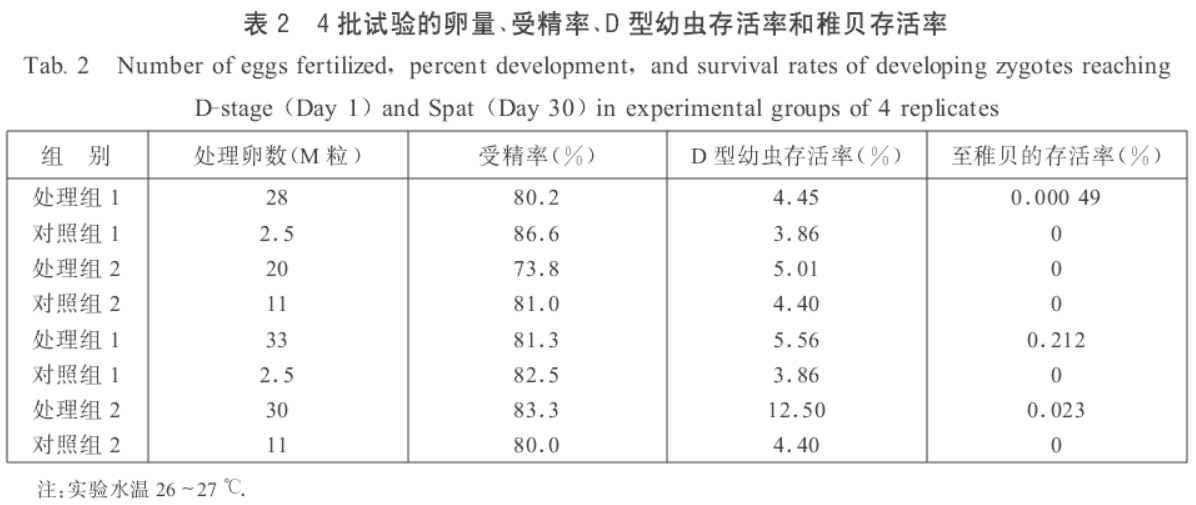
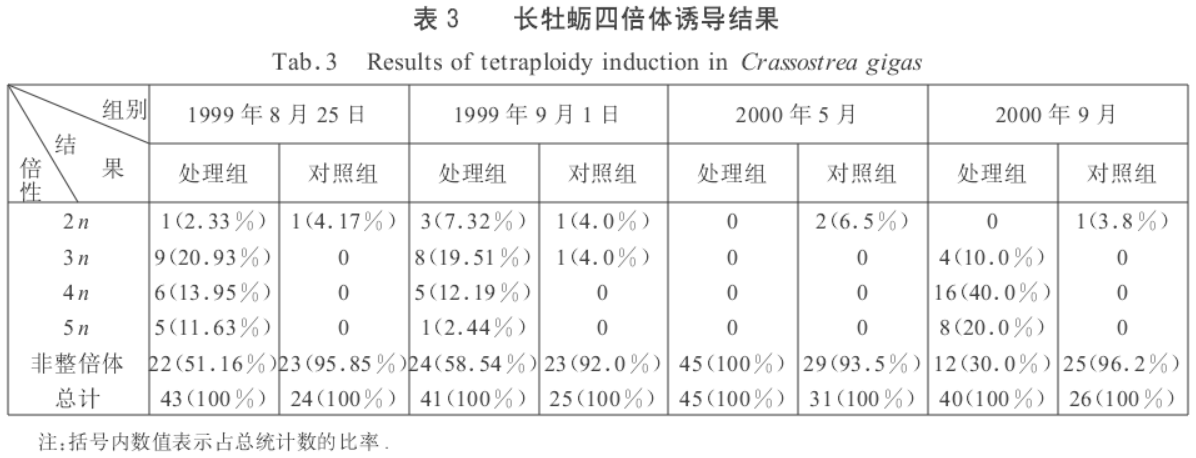
在两组处理组中诱导培育出了四倍体,处理组1最后仅获得108粒稚贝(壳长1mm),处理组4获得了7000粒的稚贝(壳长2、3mm),两个处理组的存活率分别为0.49×10- 5和23× 10- 5,比Guo和Allen(1994)报道的73.9× 10- 5低。该实验CB处理受精卵的持续时间为25min,是由于处理时间过长,导致四倍体幼虫死亡率高,抑或因为三倍体卵子成熟度或培育条件的不同影响还有待于进一步研究。处理组3诱导结果未得到四倍体,所得仅100%的非整倍体,染色体数集中在24、25条,但是幼虫存活率仍然很高,并在第9天就开始出现眼点,最后得到7万粒附着稚贝(壳长2~3mm),采用流式细胞法检测稚贝倍性,结果与担轮幼虫阶段的染色体检查结果相符,证明了染色体数在24、25条的非整倍体牡蛎也能存活,这与前人报道的结果有所不同。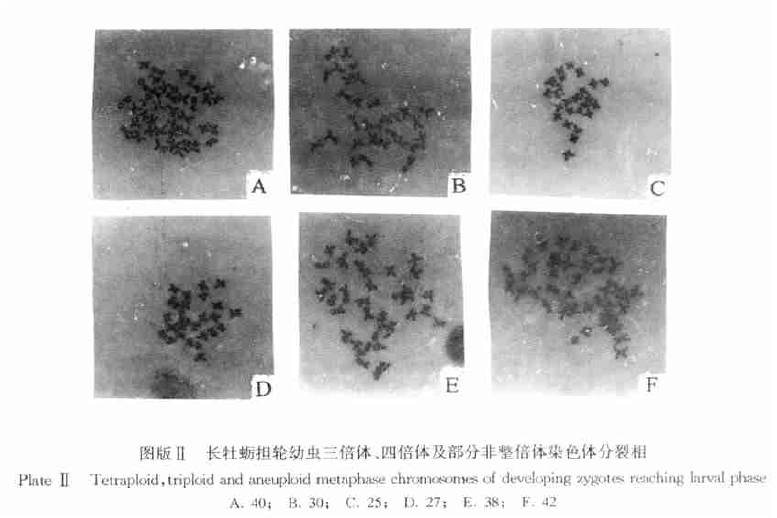
参考资料:


雌性三倍体太平洋牡蛎减数分裂中期染色体观察(延伸阅读)

卵细胞制片样品的制备 将牡蛎解剖,经性别鉴定后,挑选成熟度较好的个体,解剖取卵,卵子先后经200目和500目筛绢网过滤去除较大的组织碎片、组织液和碎屑。洗好的卵置于24℃的海水中促熟40min使其胚泡破裂,待卵子沉淀后,去掉海水,加 0.075mol/l 的氯化钾低渗40min,再用卡诺固定液(甲醇:冰 醋酸=3:1)固定,此后每隔20min,20min,40min,40min更换固定液一次,最后置4℃冰箱中保存备用。
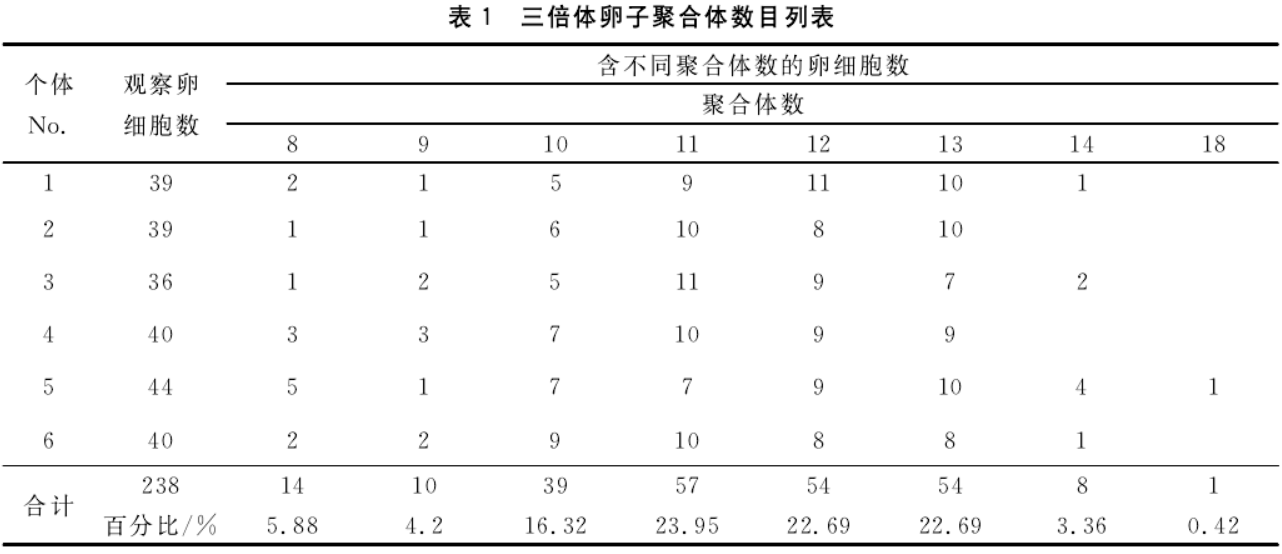
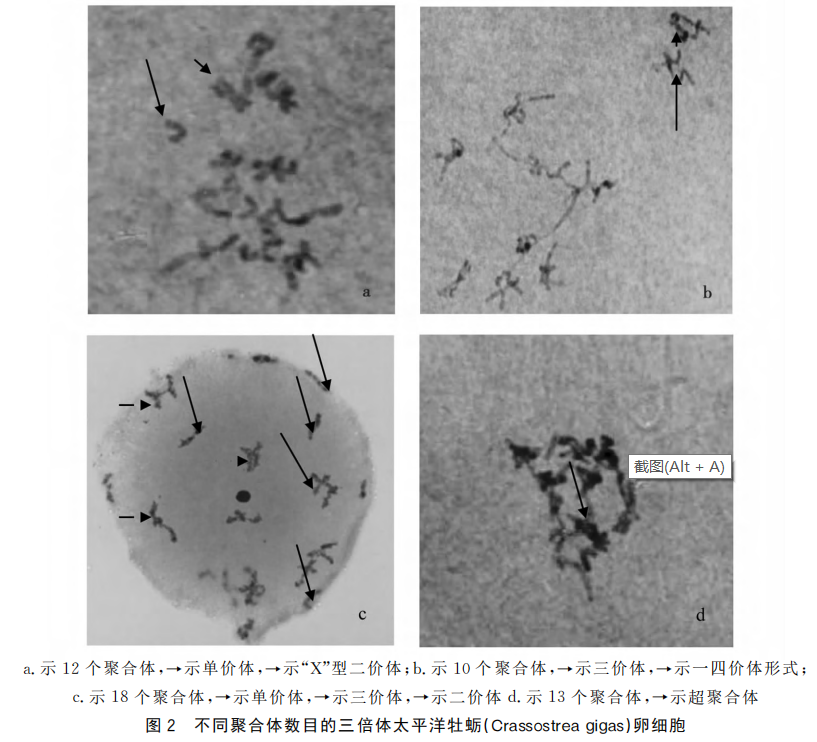
在所观察的所有细胞中,随机挑选11个染色体形态较为清晰的分裂相进行价态分析。在123个聚合体的统计观察表明三价体占总数的70.73%,其次是单价体和二价体,两者所占比例基本相当,最少的是超聚合体,仅占3.25%。
三倍体卵子发育状况:在人工诱导的贝类三倍体中,生殖腺的不育性程度很高。国内外学者曾用组织学的方法对三倍体长牡蛎生殖腺发育进行过研究,发现虽然三倍体配子发育受阻,但雌性和雄性三倍体均发育分化出一些性腺组织,仍能形成一定量的 配子。据该实验观察,三倍体卵子的发育同二倍体卵子发育相同,停滞在第一次减数分裂前期,发育较好,经24℃海水促熟部分卵子可达到第一次减数分裂中期。中期同样也有核仁的存在,而事实也证明三倍体卵子完全有能力受精。
同源染色体的配对:三倍体卵子中有三套同源染色体,共30条染色体,原则上每个同源组的三条有可能出现以下三种情况:一是形成10个二价体,10个单价体;二是10个三价体;三是单价体、二价体、三价体甚至更高多价体同时并存。在三倍体减数分裂同源染色体配对中,单价体和二价体是联会不完全的标志,三价体数目的减少就意味着单价体和二价体的增多。此外,在三倍体中存在的超聚合现象,有可能存在非同源配对现象。
同源染色体交叉与分离:在三倍体太平洋牡蛎中,这种复杂的联会形式有可能直接影响以后减数分裂染色体分离的形式。据观察,染色体分离似乎与联会程度没有关系,大多数卵子能完成两次减数分裂,类似于二倍体卵子,只是三倍体中多出的一组染色体随机分配到细胞两端。三倍体所产生的绝大多数配子的染色体数目在n和2n之间。

(2)四倍体与二倍体杂交生产三倍体



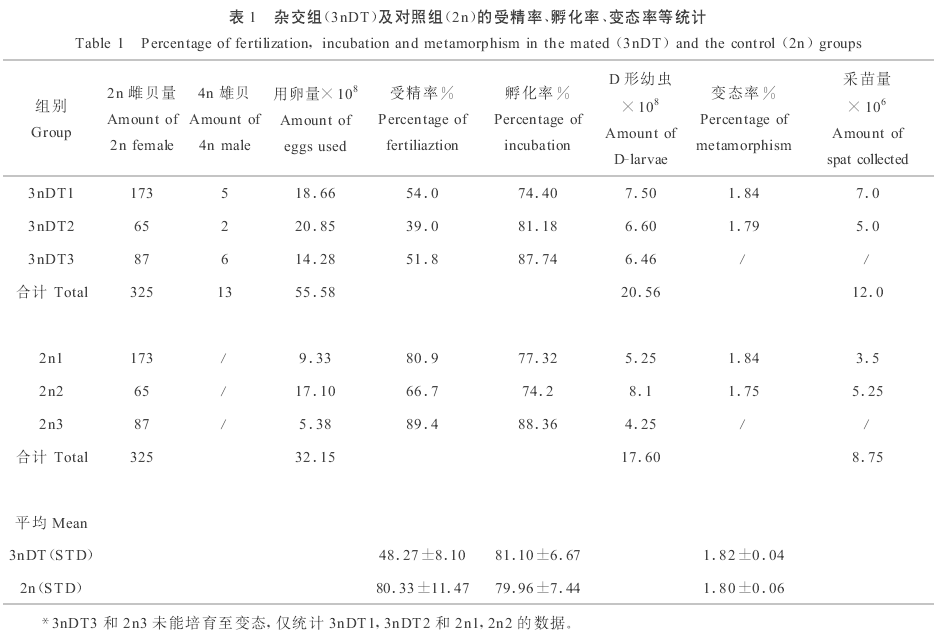
结果:
利用引进美国四倍体太平洋牡蛎(Crassostrea gigas)雄贝与我国二倍体雌贝杂交,规模化培育全三倍体牡蛎苗种。杂交组平均受精率为48.27%,对照组为80.53%,杂交组较低的受精率可能与四倍体雄贝的精子活力及精子数量有关。杂交组的孵化率、幼虫成活率和变态率分别为81.10%,42.3%和1.82%,对照组分别为79.96%,37.0%和1.80%。杂交组共培育出稚贝1200万粒。不同时期取样的流式细胞仪分析结果表明,杂交组的三倍体率为100%。
复习题:
一、问答题:
1.介绍多倍体鉴定的各种方法。
2.试述美国牡蛎三倍体研究情况。
3.试述由三倍体诱导四倍体太平洋牡蛎的方法。
二、简答题:
1.多倍体有什么生物学特性?
2.多倍体有什么用途?
3.说明由二倍体直接诱导四倍体的方法及其存在的问题。
4.介绍太平洋牡蛎四倍体与二倍体杂交生产三倍体的方法。

