

第六章 多倍体育种
第22课时 多倍体的诱导
一、自然界中的多倍体 (理解)
1.多倍体的概念和种类
(1)染色体组或基因组(genome):生物在有性繁殖过程中,或者是生物体世代交替中,能维持配子正常功能的、最低数目的
一套染色体(chromosome),称为染色体组或基因组。
(2)二倍体(diploid):两个配子结合以后形成的生物体即具有了两套染色体,称为二倍体。
(3)同源染色体:在二倍体生物的体细胞中,来自(种内)父
本和母本的两个染色体组,称为同源染色体。
4多倍体:指生物体体细胞中含有3个或3个以上染色体组
的个体。
2.自然界多倍体现象
多倍体在植物界很普遍,被子植物中大约有一半的物种是多倍体;
陆地动物中的多倍体现象比较少,但水产动物多倍体现象比较普遍,如:鲑科的虹鳟、白鲑、各种大麻哈鱼和茴鱼存在四倍体;胭脂鱼科中几乎所有的种类都是四倍体;鲫的变种-金鱼中除了四倍体外,还有三倍体如银金鱼;泥鳅以二倍体和四倍体居多。
3.天然多倍体形成的原因
(1)体细胞染色体加倍
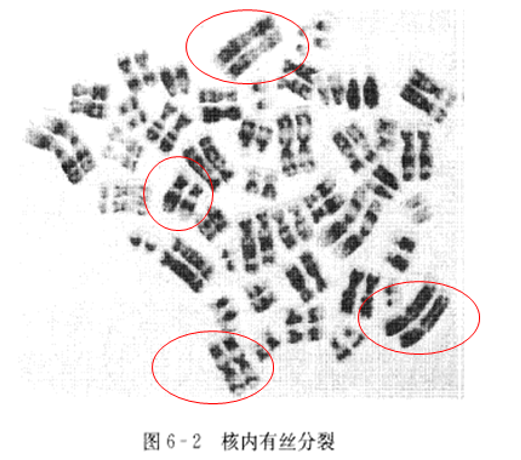
核内有丝分裂(endomitosis,核内复制):是体细胞在正常有丝分裂中,染色体复制一次,但至分裂中期,核膜没有破裂消失,也无纺锤丝的形成,因此每对染色体形成4条染色体,称为双倍染色体(diplochromosome),这时染色体两两平行排列在一起(图6-2),结果形成的细胞是四倍体。其后经过正常的细胞分裂,形成两个子细胞均为四倍体。
(2)生殖细胞的异常减数分裂
由于某种原因同源染色体或姐妹染色单体在后期没有分开一组染色体进人到同一配子中,导致配子中染色体数目的加倍形成2n 配子。这样的配子经受精后形成倍性不同于亲代的多倍体,如三倍体,但其子代的可育性很低。
(3)双(多)精受精
动物受精机制保证只有一个精子进人卵子,在一些鱼类中:精子→雄配子素Ⅱ→溶解卵膜孔的卵膜形成受精孔→卵子的雌配子素I→吸引和加速精子人卵→雌配子素Ⅱ→卵膜孔封闭→破坏孔外精子→单精受精→精卵两核的融合。
但当受精孔由于某些生理的或自然的原因不能及时封闭时,则可能有两个或多个精子入卵,形成三倍体(双精)、四倍体(三精)的合子并发育形成多倍体个体。
(4)远缘杂交
有时远缘杂交可以导致远缘多倍体的形成,这一现象在鱼类等水产动物的远缘杂交中经常出现,尤其是辅以一定的技术手段处理后更加容易获得异源多倍体。
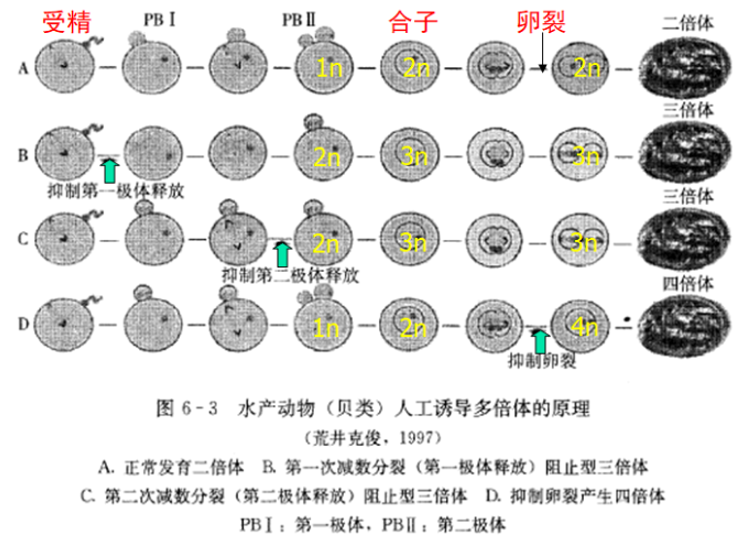
二、人工诱导多倍体的原理(掌握)
1.抑制第一极体的释放
减分I型(MI)三倍体:通过抑制第一次减数分裂诱导的三倍体。如在美洲牡蛎、太平洋牡蛎、紫贻贝、皱纹盘鲍和马氏珠母贝等多和贝类中都获得了此和三倍体。
缺点:抑制第一极体产生三倍体的方法其技术还未完全成
熟,致使诱导产生的三倍体的胚胎的孵化率和幼虫成活率都很低,不适合于大规模的商业生产。
优点:Stanley等(1984)通过对美洲牡蛎多倍体的研究发
现,MI三倍体的生长速度明显高于MII三倍体。为什么?
2.抑制第二极体的释放
减分II型(MII)三倍体:通过第二极体受阻形成的三倍体。
优点:通过抑制第二极体排出诱导多倍体(三倍体)是较为成型的技术,被广泛地应用于多种水产动物的三倍体诱导之中。
缺点:生长速度比不上MI。
3.抑制第一次卵裂
在第一卵裂期间阻止纺锤丝的形成,使卵细胞的第一次有丝分裂成为核内有丝分裂,其结果是染色体复制,一个细胞内含双倍的二倍体染色体,由此发育为一个个体即为四倍体(图6-3D)。抑制第一次卵裂的发生是目前诱导水产动物四倍体最为经济有效的方法。
4.细胞融合
细胞融合技术是用自然的或人为的方法,使种类不同的两种生物细胞直接融合,产生能够同时表达二者有益性状新杂种细胞的技术。自然进行的细胞融合,即卵子和精子受精形成合体,只能发生在同一种生物或亲缘关系很近的生物种之间。然而,细胞融合技术的问世不仅能产生同种细胞融合,也能产生种间细胞的融合,不仅为细胞学、遗传学的研究提供了有力的手段,而且开创了动物、植物育种和工业生产的新局面。
三、人工诱导多倍体的方法 (掌握)
(一) 物理学方法
1.温度休克法:
(1)冷休克法
低温能阻止纺锤丝形成,细胞不能正常分裂,可获得二倍体的卵核,受精即可获得三倍体。
无论抑制哪一个极体的排出,均需要在减数分裂的中期,即纺锤丝形成前和形成期间进行。
冷休克处理的温度原则是略高于致死温度。
(2)热休克法:
高温环境中,纺锤体供能途径受阻,细胞分裂被抑制,极体不能排放。
热休克处理的时间必须刚好在减数分裂的中期以前及中期分裂纺锤丝形成期间。
对于贝类可以是第一次减数分裂,也可以是第二次减数分裂;但对于鱼类目前仅对第二次减数分裂有效。如果诱导休克发生在第一次卵裂,则形成四倍体。
喜冷性鱼类宜用热休克;温水性鱼类宜用冷休克:
温度范围以不致死鱼卵或少量致死为度;
进行温度处理最重要的是必须确定处理的开始时间、持续时间以及温度高低。
2.静水压处理
将处于第一或第二成熟分裂期的受精卵采用较高的静水压处
理(大约为650kgf/cm2),破坏纺锤丝的形成。
3.电脉冲休克
在一定的电场条件下,细胞与细胞之间的膜会发生融合,导致两个细胞合二而一,从而诱发多倍体(四倍体)。例如,将银大麻哈鱼受精40min后的卵置于26℃,并给以10min的交流电脉冲
休克处理可得到100%的三倍体。
(二)化学方法
1.细胞松弛素B(CB)
真菌的一类代谢产物,特异性破坏微丝,抑制细胞质分裂,阻止极体的释放,从而产生多倍体。
CB是一种致癌物质,水溶性较差,必需将其溶解在1%的二甲亚砜(DMSO)中,配制成一定浓度的药液,处理结束后需用含1%DMSO的溶液浸洗受精卵,去除残留的CB。
例如,在25℃条件下,用1mg/L的CB处理太平洋牡蛎受精卵(受精后30~45min)15min,诱发得到88%的三倍体。
2.6-二甲基氨基嘌呤(简称6-DMAP)
是嘌呤霉素的一种类似物,破坏微管的聚合中心,抑制卵细胞极体的形成和释放。
6-DMAP非致癌性,具水活性。洗除该药物后,受精卵可以正常发育。
6-DMAP低毒、高效、价格便宜,在诱导多倍体方面表现出较大的优越性和较广阔的应用前景。
3.聚乙二醇(PEG)
是一种常用的细胞融合剂,能使细胞发生融合,从而使染色体加倍。该药物被广泛用于试管内的细胞融合。由于其特殊的理化特性,也将其用于水产动物的多倍体诱导。Ueda等(1986)用聚乙二醇处理虹鳟精子,然后受精,获得了2/5的三倍体胚胎,经染色体组型分析,两套染色体来自父本,一套来自母本。证明三倍体的形成是由融合的双精子受精所致。
4.咖啡因(caffeine)
咖啡因进人细胞后可以立即提高细胞内的Ca2+浓度。由于微管对Ca2+浓度变化敏感,当Ca2+浓度极低或高于1mg/mL时,会引起微管二聚体的解聚,阻止细胞分裂,从而形成多倍体。
(三)生物学方法
1.远缘杂交
亲本配子染色体加倍可以使染色体达到平衡,从而克服远缘杂交的困难,获得多倍体的杂交种。因此,远缘杂交可以产生异源多倍体。
2.核移植及细胞融合
核移植诱导鱼类多倍体技术仍处于实验阶段,试管内的细胞
融合可以产生多倍体细胞。
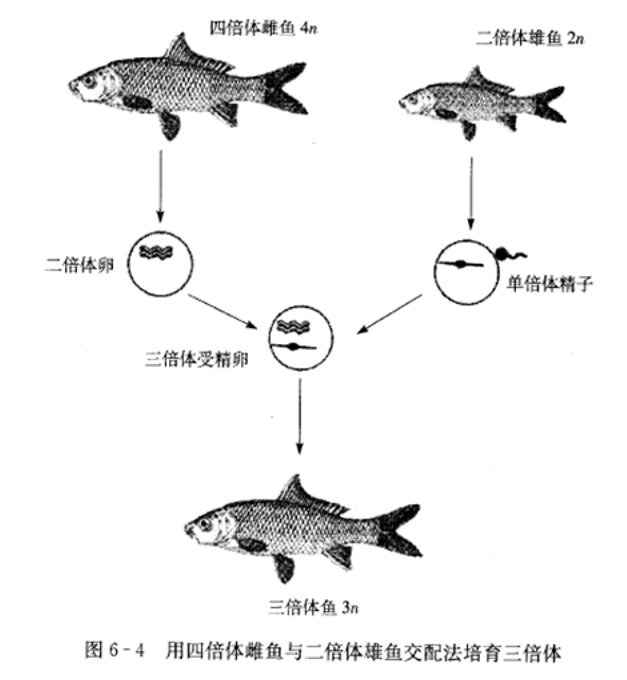
3.四倍体与二倍体交配生产三倍体
(1)虹鳟受精卵用CB处理,以及用热休克和静水压处理都获得了四倍体。已用四倍体商业化生产虹鳟三倍体苗种。
(2)刘筠等用鲫和鲤交杂获得四倍体鲫鲤,然后将其与鲤或鲫二倍体回交获得了三倍体的杂种,分别称为湘云鲤和湘云鲫。另外,已经获得奥利亚罗非鱼的四倍体个体、兴国红鲤与草鱼异源四倍体、鲢的四倍体。
(3)2004年专家对中国海洋大学培育的四倍体牡蛎进行活体倍性检测后,一致认为海大培养存活的10月龄的四倍体牡蛎培育取得成功。据了解,四倍体牡蛎群成活率高达93%,这是多年来我国贝类四倍体育种研究取得的首次成功,也打破了少数发达国家的技术垄断。
http://news.sina.com.cn/o/2004-06-20/16012857913s.shtml
(4)利用引进四倍体培育全三倍体太平洋牡蛎苗种
王昭萍,姜波,孔令峰,于瑞海,王如才,郭希明
《中国海洋大学学报(自然科学版)》 2004年05期
http://www.cnki.com.cn/Article/CJFDTotal-QDHY200405008.htm
利用引进美国的四倍体太平洋牡蛎(Crassostrea gigas)雄贝与我国的二倍体雌贝杂交,规模化培育全三倍体牡蛎苗种。杂交组的平均受精率为 48.27%,对照组为80.53%,杂交组较低的受精率可能与四倍体雄贝的精子活力及精子数量有关。杂交组的孵化率、幼虫成活率和变态率分别为 81.10%、42.3 %和1.82%,对照组分别为79.96%、37.0%和1.80%。杂交组共培育出稚贝1200万粒。不同时期取样的流式细胞仪分析结果表明,杂交组的三倍体率为100%。

四、诱导多倍体的细胞学特征(掌握)
1.抑制第一极体释放的染色体分离方式:
Guo等(1994)在太平洋牡蛎(2n=20)受精卵的细胞学观察研究中发现,经CB处理抑制第一极体释放后,染色体产生正常和非正常的分离,前者形成正常的三倍体,称为MI 三倍体,但仅占7%;后者大部分形成非整倍体2.4n/2.5n/3.7和少量的整倍体2n/3n/4n。
在第二次减数分裂中,染色体的分离出现以下几种方式:
①联合二极分离(正常的分离):
细胞核内形成二倍体的雌性原核与正常的雄性原核结合就形成三倍体。
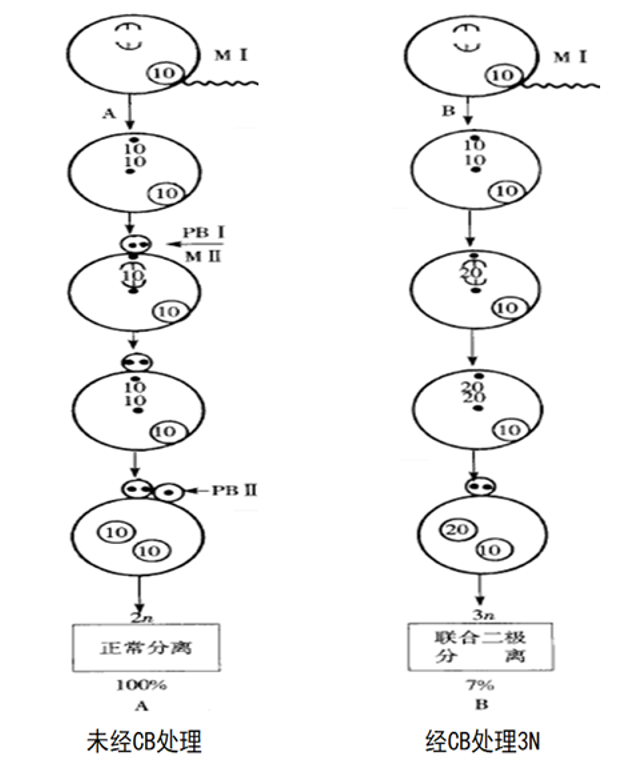
②随机三极分离(非正常的分离):下图
③非混合三极分离(非正常的分离):
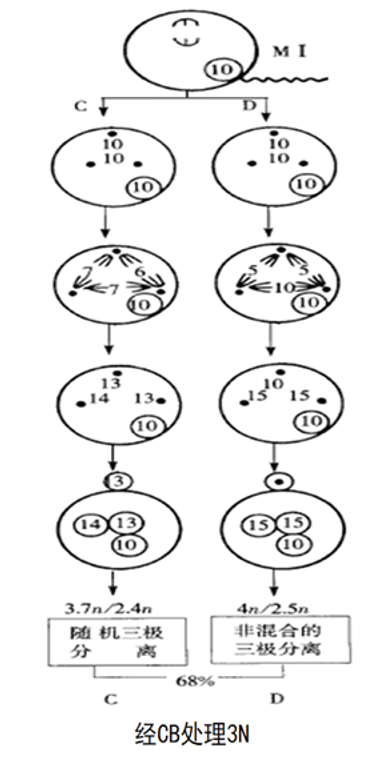
④独立二极分离(非正常的分离):
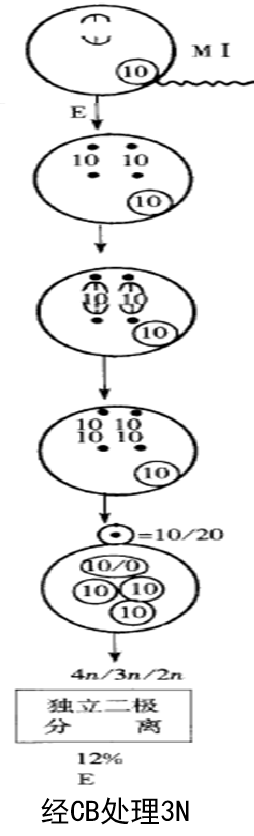
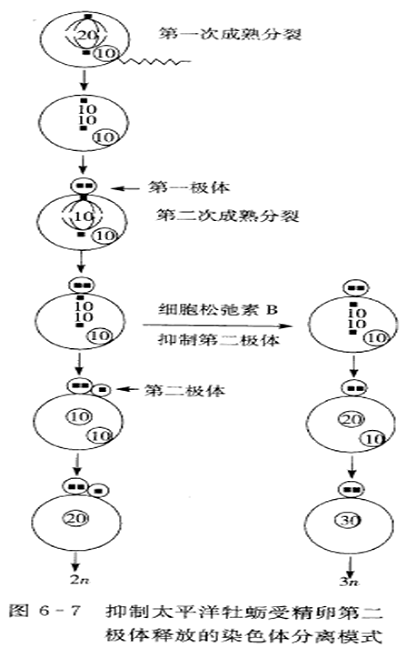
2.抑制第二极体释放的染色体分离方式
抑制受精卵第二极体释放形成的三倍体称为MII 三倍体,第二极体释放的染色体分离方式因种类不同而不同,一般成为整三倍体。以太平洋牡蛎为例说明抑制第二极体释放的遗传结果。
复习题:
一、问答题:
1.综述人工诱导多倍体的方法。
2.以太平洋牡蛎为例画图说明抑制第一极体释放的染色体分离方式。
二、简答题:
1.天然多倍体形成的原因有哪些?
2.抑制第一极体释放诱导三倍体的优缺点是什么?
3.抑制第二极体释放诱导三倍体的优缺点是什么?
4.试述人工诱导多倍体的原理。
5.人工诱导多倍体的物理学方法有哪些?
6.人工诱导多倍体的化学方法有哪些?
7.人工诱导多倍体的生物方法有哪些?
8.以太平洋牡蛎为例画图说明抑制第二极体释放的染色体分离方式。
三、名词解释:
1.染色体组;
2.二倍体;
3.同源染色体;
4.多倍体;
5.减分I型(MI)三倍体;
6.减分II型(MII)三倍体;
7.细胞融合。

