
3.5
影响光合作用的因素
5. Photosynthetic product and comparison of 3 pathways.ppt(下载附件 2.99 MB)
植物光合作用受到外界环境因素和内部因素的影响而发生变化。而要了解这些因素对光合作用影响的大小及作用机理,首先要了解光合作用的指标。
3.5.1 光合作用的指标
光合作用的指标是光合速率和光合生产率。
光合速率(photosynthetic rate)的大小可用单位时间单位叶面积所吸收的CO2或释放的O2表示。一般常用毫克CO2/分米2·小时表示,即每小时每平方分米叶面积吸收的CO2毫克数。现在,越来越多地使用单位时间单位叶面积所吸收的CO2或释放的O2微摩尔来表示,即用mmol.m-2.s-1或mmol.dm-2.h-1来表示。通常测定光合速率时,没有把叶片的呼吸作用考虑在内,即测定的光合速率等于光合作用与呼吸作用的差数,称为表观光合速率(apparent photosynthetic rate),又称净光合速率(net photosynthetic rate, Pn)。表观光合速率与呼吸速率之和称为真正光合速率(true photosynthetic rate)或叫总光合速率 (gross photosynthetic rate)。
在农业生产中衡量作物一定时间内光合作用产物的净积累量,通常用光合生产率(photosyntheticproduce rate),又称净同化率(netassimilation rate, NAR)来表示,即在较长时间内(例如一昼夜或一周)单位叶面积生产的干物质量(即较长时间内的表观光合速率)。常用g·m-2·d-1表示 。由于测定时间较长,存在着夜间的呼吸作用和光合作用产物从叶片向外运输以及用于其它代谢过程的消耗。因此,测得的光合生产率低于短期测得的光合速率。
3.5.2 外界因素对光合速率的影响
3.5.2.1光照
光是光合作用的能量来源,是叶绿素合成的必要条件,还调节碳同化循环许多酶的活性和气孔开度,因此光是影响光合作用的重要因素。
1、 光强度对光合作用的影响
(1)光强度和光合速率的关系
光合作用是一个光生物化学反应,所以光合速率随着光照强度的增加而加快。在一定范围内几乎是呈正相关。但超过一定范围之后,光合速率的增加却转慢;当达到某一光照强度时,光合速率就不再增加,这种现象称为光饱和现象(light saturation),刚出现光饱和现象时的光强度称为光饱和点(light saturation point)。同一叶子在同一时间内,光合作用过程中吸收的CO2和呼吸作用过程中放出的CO2等量时的光照强度,就称为光补偿点(light compensation point),光极弱时,即光强度低于光补偿点时,真正光合速率低于呼吸速率,呼吸放出的二氧化碳多于光合作用同化的二氧化碳,表观上叶片释放二氧化碳。在黑暗中光合作用等于零,只有呼吸作用。上述光强度与光合速率的关系可以以图3-31的曲线表示,该曲线也称需光量曲线。
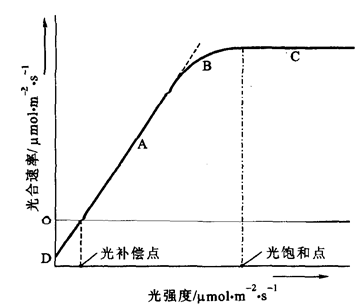
图3-31 光强度-光合速率的关系
植物出现光饱和点的实质是强光下暗反应跟不上光反应从而限制了光合速率随着光强度的增加而提高。因此,从光强度-光合曲线中可以看出影响光合速率的主要因素随光强度的不同而不同。大于光饱和点光强度下限制光合作用的主要因素有CO2扩散速率(受CO2浓度影响)和CO2固定速率(受羧化酶活性和RuBP再生速率影响)。弱光下,光强度是控制光合的主要因素,曲线的斜率即为表观量子效率。曲线的斜率大,表明植物吸收与转换光能的色素蛋白复合体可能较多,利用弱光的能力强。实测的表观量子效率一般在0.03~0.05之间。随着光强度增高,叶片吸收光能增多,光化学反应速率加快,产生的同化力多,于是CO2固定速率加快。此外,气孔开度、Rubisco活性及光呼吸速率也影响直线阶段(A)的光合速率,因为这些因素都会随光强度的提高而增大,其中气孔开度、Rubisco活性的提高对光合速率有正效应,光呼吸速率的提高对光合速率有负效应。
各种植物的光饱和点不同,与叶片厚薄、单位叶面积叶绿素含量多少有关。根据对光照强度需要的不同,可把植物分为阳生植物(sun plant)和阴生植物(shade plant)两类。阳生植物叶片的光饱和点为90~180 mmol.m-2.s-1或更高,阴生植物叶片的光饱和点为360~450 mmol.m-2.s-1。上述光饱和点的数值是指单叶而言,对群体则不适用。因为植物群体对光能的利用,与单株叶片不同。群体叶枝繁茂,当外部光照很强,达到单叶光饱和点以上时,而群体内部的光照强度仍在光饱和点以下,中、下层叶片就比较充分地利用群体中的透射光和反射光。群体对光能的利用更充分,光饱和点就会上升。
光补偿点在实践上在很大的意义。间作和套作时作物种类的搭配,林带树种的配置,间苗、修剪、采伐的程度,冬季温室栽培蔬菜等等都与补偿点有关。又如,栽培作物由于过密或肥水过多,造成徒长,封行过早,中下层叶子所受的光照往往在光补偿点以下,这些叶子不但不能制造养分,反而消耗养分形成消费器官。因此,生产上要注意合理密植,肥水管理恰当,保证透光性好。
植物的光饱和点和光补偿点都不是固定值,它们会随外界条件的变化而变动。如当CO2浓度升高时,光饱和点增加,而光补偿点降低。
(2)光合作用的光抑制
光是光合作用的能源,所以光是光合作用必需的。然而,光照过强时,当植物吸收的光能超过其所需要时,过剩的光能会导致光合速率下降,这种现象称为光合作用的光抑制(photoinhition of photosynthesis)。
关于光抑制的机制,一般认为光抑制主要发生在PSⅡ。在特殊情况下,如低温弱光也会导致PSⅠ发生光抑制。正常情况下,光反应与暗反应协调进行,光反应中形成的同化力在暗反应中被及时用掉。但由于叶绿体基质中的CO2浓度往往很低,接近CO2补偿点,当光照过强时,光能过剩。一方面因NADP+不足使电子传递给O2形成超氧阴离子自由基(O2-);另一方面会导致还原态电子的积累,形成三线态叶绿素(chlT),chlT与分子氧反应生成单线态氧(1O2)。O2-和1O2都是化学性质非常活泼的活性氧,如不及时清除,它们会攻击叶绿素和PSⅡ反应中心的D1蛋白,从而损伤光合机构。如果强光时间过长,甚至会出现光氧化现象,光合色素和光合膜结构遭受破坏。如果植物遭受低温、高温、干旱等不良环境因子胁迫的同时,又遇到高光强会加剧光抑制的危害。例如,黄瓜等对冷害敏感的植物,在暗中受冷不会影响光合作用,但在光和低温下,则光合磷酸化受抑制,细胞膜透性加大。
植物在长期的进化过程中也形成了多种光保护机制。①细胞中存在着活性氧清除系统,如超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、谷胱甘肽、抗坏血酸及类胡萝卜素等,它们共同防御活性氧对细胞的伤害;②通过代谢耗能,如提高光合速率,增强光呼吸和Mehler反应等;③提高热耗散能力,如依赖叶黄素循环的非辐射能量耗散;④PSⅡ的可逆失活与修复。植物本身对光抑制有一定程度的保护性反应。例如,叶子运动,调节角度去回避强光;叶绿体运动以适应光照强弱。又如,小麦幼苗在强光下,叶绿体中的捕光叶绿素a/b蛋白复合体含量低于生长在弱光下的;而负担将光能转化为化学能的反应中心复合体含量在强光下大于在弱光下。在农业生产上,要尽可能提供作物生长发育所需的条件,尤其是要防止几种胁迫因子同时出现,最大限度地减轻光抑制。
2、光质对光合作用的影响
如前所述,太阳辐射中只有可见光部分才能被植物光合作用利用。但由于植物的种类不同、生长发育的环境不同及生长发育的时期不同,光合作用色素的种类、含量和比例也不同。因此,光质不同也影响植物的光合速率。菜豆在橙、红光下光合速率最快,蓝、紫光其次,绿光最差。其他高等植物和绿藻都有类似结果。
一般来说,不同光波影响下的光合高峰相当于叶绿素和类胡萝卜素的吸收光谱高峰。在自然条件下,植物会或多或少受到不同波长的光线照射。例如,阴天的光照不仅光强减弱,而且蓝光和绿光增多;树木的叶片吸收红光和蓝光较多,故树冠下的光线富于绿光,尤其是树木繁茂的森林更是明显。水层同样改变光强和光质。水层越深,光照越弱,例如,20m深处的光强和水面的光强比较,前者为后者的二十分之一。水层对光波中的红、橙部分吸收显著多于对蓝、绿部分的吸收,水下深层的光线相对富于短波长的光。所以含有叶绿素,吸收红光较多的绿藻分布海水的表层,含有藻红蛋白,吸收绿、蓝光较多的红藻,则分布在海水的深层。这是海藻对光线的适应。
3.5.2.2 CO2对光合作用的影响
二氧化碳是光合作用的原料,对光合速率的影响很大。
由图3-32可以看出, 在光下CO2浓度等于零时,光合作用器官只有呼吸作用释放CO2。随着CO2浓度的增加光合速率增加,当光合作用吸收的CO2等于呼吸作用放出的CO2量时,即光合速率与呼吸速率相等时,外界的CO2浓度叫做CO2补偿点(CO2 compensationpoint,图中的C点)。尔后随着CO2浓度提高,光合速率直线增加。但是随着CO2浓度的进一步增加,光合速率变慢,当CO2浓度达到某一范围时,光合速率达到最大值(Pm),光合速率开始达到最大值时的CO2浓度被称为CO2饱和点(CO2 saturation point,图中的Pm点)。
在低CO2浓度条件下,CO2浓度是光合作用的限制因子,直线的斜率(CE)受羧化酶活性和量的限制。因而,CE被称为羧化效率。CE值大,则表示在较低的CO2浓度下有较高的光合速率,亦即Rubisco的羧化效率较高。

图3-32 CO2 ¾光合作用曲线模式图
在CO2浓度达到CO2饱和点以上,CO2已不再是光合作用的限制因子,而CO2受体的量,即RuBP的再生速率成了影响光合的因素。由于RuBP的再生受同化力供应的影响,所以饱和阶段的光合速率反映了光反应活性,即光合电子传递和光合磷酸化活性,因而Pm被称为光合能力。
水稻单叶的CO2补偿点是55 mg/LCO2(25℃,光照>10klx),其变化范围随光照强度而异。光照弱时,光合速率降低比呼吸速率显著,所以要求较高的CO2水平,才能维持光合与呼吸相等,也即是CO2补偿点高。光照强时,光合显著大于呼吸,CO2补偿点就低。作物高产栽培的密度大,肥水充足,植株繁茂,吸收更多的CO2,特别是在中午前后,CO2就成为增产的限制因子之一。
陆生植物光合作用所需要的碳源,主要是空气中的CO2,CO2主要是通过叶片气孔进入叶子。空气中的CO2含量一般占体积的0.033%(即0.65mg/L,0℃,101 kPa),对植物的光合作用来说是比较低的。光合过程中吸收相当大量的CO2,如向日葵叶面积吸收0.14CO2cm3/h·cm2。气孔在叶面上所占的面积不到1%,但由于小孔扩散的特点。空气中的二氧化碳经过气孔进入叶肉细胞的细胞间隙,是以气体状态扩散进行的,速度很快;但当二氧碳经过胞壁透到叶绿体时,便必须溶解在水中,扩散速度也大减。
陆生植物的根部也可以吸收土壤中的CO2和碳酸盐,用于光合作用。试验证明,把菜豆幼苗根部放在含的14CO2的空气中或NaH14CO3的营养液中,进行光照,结果光合产物中都发现14C。关于根部吸收的CO2如何用于光合作用问题,可能是CO2进入根后就与丙酮酸结合成草酰乙酸,再还原为苹果酸,苹果酸沿输导组织上升而进入绿色器官——叶、茎和果实中。如果这时在光照下,则用于光合作用;如果在黑暗中,大部分的CO2就排出体外。
浸没在水中的绿色植物,其光合作用的碳源是溶于水中的CO2、碳酸盐和重碳酸盐,这些物质可通过表皮细胞进入叶子中去。
增加CO2浓度固然可以提高光合速率。但是,随着世界范围内消耗石油燃料的急剧增加,排放到大气中CO2浓度的增加会产生“温室效应”(greenhouse effect)。所谓温室效应,是指在地球周围的大气层中,人类无限制地向地球大气层中排放CO2,使CO2浓度不断增长。本来太阳辐射下来的热,地球以红外线形式重新辐射到空间。由于大气层中的CO2能强烈地吸收红外线,太阳辐射的能量在大气层中就“易入难出”,温度上升,似温室一样(图3-33)。地球变暖,造成冰川融化,海水上升,会淹没沿海城市和农田;气候也异常,高温、干旱。所以温室效应已引起全球关注。防止温室效应加剧的办法是尽量减少燃烧时排放CO2,积极种植树木,吸收CO2,维持大气中CO2浓度的平衡。
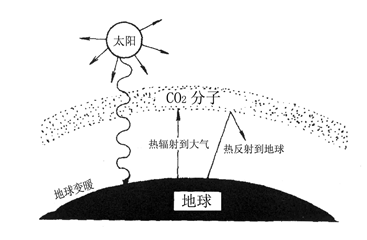
图3-33 地球大气层中CO2的温室效应
3.6.2.3温度
光合过程中的暗反应是由酶所催化的化学反应,而温度直接影响酶的活性,因此,温度对光合作用的影响也很大。除了少数的例子以外,一般植物可在10~35℃下正常发进行光合作用,其中以25~30℃最适宜,在35℃以上时光合作用就开始下降,40~50℃时即完全停止。植物光合作用温度的三基点因种类的不同而不同。一般而言,乃寒植物光合作用的最低和最适温度低于喜温植物,而最高温度相似。在低温中,酶促反应下降,故限制了光合作用的进行。光合作用在高温时降低的原因,一方面是高温破坏叶绿体和细胞质的结构,并使叶绿体的酶钝化;另一方面是在高温时,呼吸速率大于光合速率,因此,虽然真正光合作用增大,但因呼吸作用的牵制,表观光合作用便降低。
3.6.2.4 矿质元素
矿质元素直接或间接影响光合作用。氮、镁、铁、锰等是叶绿素生物合成所必需的矿质元素,钾、磷等参与糖类代谢,缺乏时便影响糖类的转变和运输,这样也就间接影响了光合作用;同时,磷也参与光合作用中间产物和能量转变,所以对光合作用影响很大;铜和铁是电子传递体的重要成分;锰、氯和钙是OEC的必需因子;钾是气孔开关的重要因子。三要素中以氮肥对光合作用的效果最明显。追施氮肥促使光合速率的原因有两方面:一方面是促进中叶片面积增大,叶片数目增多,增加光合面积,这是间接的影响。另一方面是直接的影响,即影响光合能力。施氮肥后,叶绿素含量急剧增加,加速光反应;氮肥亦增加叶片蛋白态氮百分率,而蛋白质是酶的主要组成成分,也使暗反应进行顺利。但是,农业生产中要合理施肥才能使光合作用调节到最适状态,达到增产的目的。
3.5.2.5水分
水分是光合作用原料之一,缺乏时可使光合速率下降。水分在植物体内的功能是多方面的,叶子要在含水量较高的条件下才能生存,而光合作用所需的水分只是植物所吸收水分的一小部分(1%以下)。因此,水分缺乏主要是间接的影响光合作用下降。具体来说,缺水使气孔关闭,影响二氧化碳进入叶内;缺水使叶片淀粉水解加强,糖类堆积,光合产物输出缓慢,反馈抑制光合作用;缺水抑制叶片生长,光合面积下降;缺水严重时叶绿体及片层结构破坏。这些都会使光合速率下降。试验证明,由于土壤干旱而处于永久萎蔫的甘蔗叶片,其光合速率比原来正常的下降87%。再灌以水,叶子在数小时后可恢复膨胀状态,可是表观光合速率在好几天后仍未恢复正常。由此可见,叶片缺水过甚,会严重损害光合进程。水稻烤田,棉花、花生炼苗时,要认真控制烤田(炼苗)程度,不能过头。
3.5.2.6氧
实验证明,当将环境的氧含量降低为1~3%时,就发现正常大气中21%氧含量对植物光合作用是有抑制效应的,通称之为氧的胁迫。大气中21%氧含量对C3植物的光合作用抑制竟达33~50%之高,而对C4植物几乎不抑制。氧抑制光合作用的原因主要是:氧分压增加提高了Rubisco加氧酶活性,提高光呼吸速率;氧能与NADP+竞争接受电子,NADPH合成量就少,碳同化所需的还原能力减少;氧接受电子后形成的超氧自由基,会破坏光合膜;在强光下,氧参与光合色素的光氧化,破坏光合色素等等。但是,植物在长期演化过程中逐渐形成一些保护性机制,例如,叶绿体中生成的超氧物歧化酶可以消除超氧自由基,抗坏血酸过氧化物酶可以消除过氧化氢等。在农业生产上喷施150mg/L2,3-环氧丙酸,可部分拮抗氧抑制,提高光合速率。
3.5.2.7光合速率的日变化
影响光合作用的外界条件每天都在时时刻刻变化着,所以光合速率在一天中也有变化。在温暖的日子里,如水分供应充足,太阳光照成为主要矛盾,光合过程一般与太阳辐射进程相符合:从早晨开始,光合作用逐渐加强,中午达到高峰,以后逐渐降低,到日落则停止,成为单峰曲线。这是指无云的晴天而言。如果白天云量变化不定,则光合速率随着到达地面的光强度的变化而变化,成不规则的曲线。但当晴天无云而太阳光照强烈时,光合进程便形成双峰曲线:一个高峰在上午,一个高峰在下午。中午前后光合速率下降,呈现“午休”(midday depression of photosynthesis)现象。为什么会出现这种现象呢?(1)水分在中午供给不上,气孔关闭;(2)CO2供应不足;(3)光呼吸增加。这些都表现为光合作用速率的下降。南方夏季日照强,作物“午休”会更普遍一些,在生产上应适时灌溉或选用抗旱品种,以缓和“午休”现象,增加光合能力。
3.5.3 内部因素对光合速率的影响
不同植物(C3植物、C4植物、CAM、阳生植物和阴生植物等)、相同植物不同部位及相同部位的不同发育阶段光合作用速率都不同。
3.5.3.1 不同部位
由于叶绿素具有接受和转换能量的作用,所以,植株中凡是绿色的、具有叶绿素的部位都进行光合作用,在一定范围内,叶绿素含量越多,光合越强。就拿抽穗后的水稻植株来说,叶片、叶鞘、穗轴、节间和颖壳等部分都能进行光合作用。但一般而言,叶片光合速率最大,叶鞘次之,穗轴和节间很小,颖壳甚微。在生产上尽量保持足够的叶片,制造更多光合产物,为高产提供物质基础。
以一片叶子来说,最幼嫩的叶片光合速率低,随着叶子成长,光合速率不断加强,达到高峰,后来叶子衰老,光合速率就下降。根据这个原则,同一植株不同部位的叶片光合速率,因叶子发育状况不同而呈规律性的变化。小麦和水稻等靠近穗的旗叶光合速率最高,光合作用时间也最长。
3.5.3.2 不同生育期
一株作物不同生育期的光合速率,一般都以营养生长中期为最强,到生长末期就下降。就拿水稻来说,分蘖盛期的光合速率最快,以后随生育期的进展而下降,特别在抽穗期以后下降较快。但从群体来看,群体的光合量不仅决定于单位叶面积的光合速率,而且很大程度上受总叶面积及群体结构的影响。水稻群体光合量有两个高峰:一个在分蘖盛期,另一个在孕穗期。从此以后,下层叶片枯黄,单株叶面积减少,因此光合量急剧下降。在农业生产上,通过栽培措施以延长生育后期的叶片寿命和光合功能,使生育后期光合下降缓和一些,更有利于种子饱满充实。

