
3.4 光呼吸
4photorespiration.ppt(下载附件 673 KB)
中文课件:
3.4.1 光呼吸的现象与定义
光呼吸(photorespiration)是指植物绿色组织在光下与光合作用相联系而发生的吸收氧和释放CO2的过程,也称为“氧化的光合碳循环”,简称为C2循环。
光呼吸与一般呼吸(暗呼吸)不同的是:光呼吸只在光下进行,而暗呼吸则在光下或黑暗中均可进行;光呼吸氧化的底物是乙醇酸,暗呼吸氧化的底物通常是糖等。此外,催化这两个过程的酶和它们在细胞内进行的部位也不同。还需要指出,绿色组织在光下的吸氧不全是由于光呼吸,其他耗氧反应如光合作用的Mehler反应,以及线粒体在光下进行的暗呼吸氧化反应等,也包括在光下吸氧范畴之内。
光呼吸的发现,最初可以追溯到1920年Warburg报告氧对小球藻光合作用的抑制(称Warburg effect)。其后Decker在1955年观察到在停止光照后, 烟草叶片大量放出CO2的过程,称为“CO2猝发”(CO2 outburst),认为是光下呼吸释放CO2的延续,即在光下所形成的光呼吸底物尚未立即用完,在停止光照后光呼吸底物的继续氧化。Zelitch在1964年证明乙醇酸是光呼吸的底物。Tolbert在1971年提出一个完整的光呼吸代谢途径。这些研究加上其他人的研究,便确定了光呼吸的代谢过程。
3.4.2 光呼吸的生物化学过程
光呼吸是一个氧化过程,涉及到叶绿体、过氧化物体和线粒体三种细胞器。被氧化的底物是乙醇酸。植物的绿色组织要在光照下(黑暗不行)才能形成乙醇酸(glycolic acid)。因为乙醇酸首先由Rubisco催化RuBP产生磷酸乙醇酸(phosphoglycolicacid),后者在磷酸酶作用下,脱去磷酸而产生乙醇酸。这些过程是在叶绿体内进行的。
乙醇酸形成后就转移到过氧化物体(peroxisome)。过氧化物体是一种细胞器,直径为0.2~1.5μm,只有单层膜。所有高等植物的光合细胞中均有过氧化物体。C3植物叶肉细胞的过氧化物体较多,而C4植物的过氧化物体大多数在维管束鞘细胞的薄壁细胞内。在过氧化物体内,乙醇酸在乙醇酸氧化酶(glycollic acid oxidase或glycolate oxidase)作用下,被氧化为乙醛酸(glyoxlate或glyoxylic acid)和过氧化氢。过氧化氢在过氧过氢酶的作用下分解,产生水并放出氧气。乙醛酸在转氨酶作用下,从谷氨酸得到氨基而形成甘氨酸。
甘氨酸的进一步转化是在线粒体中进行的。两个分子甘氨酸转发生氧化脱羧和羟甲基转移反应变为丝氨酸并释放CO2。丝氨酸再进入过氧化物体,经转氨酶的催化,形成羟基丙酮酸。羟基丙酮酸在甘油酸脱氢酶作用下,还原为甘油酸。最后,甘油酸在叶绿体内经过甘油酸激酶的磷酸化,产生3-磷酸甘油酸,参加卡尔文循环的代谢。进一步由核酮糖二磷酸形成乙醇酸。乙醇酸途径到此结束(图3-28)。在整个乙醇酸途径中,O2的吸收发生于叶绿体(反应②)和过氧化物体(反应③),CO2的放出发生于线粒体(反应⑥)中。由于光呼吸的底物乙醇酸是C2化合物,其氧化产物乙醛酸以及其转氨形成的甘氨酸都是C2化合物,故也称这条途径为C2光呼吸碳氧化环 [C2 photorespiratorycarbonoxidation(PCO) cycle],简称C2环。
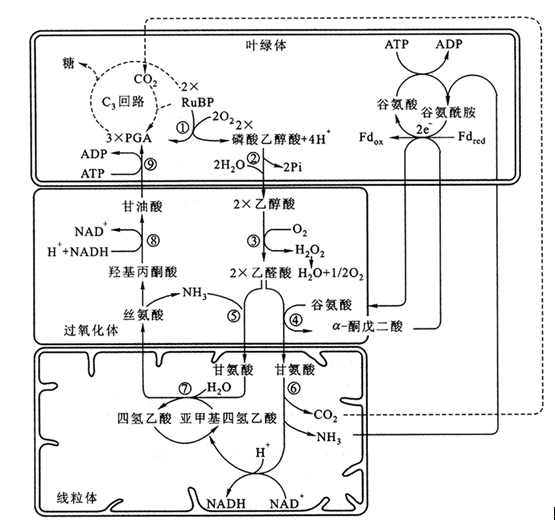
图3-28 光呼吸生物化学过程及涉及的细胞器
①Rubisco;②磷酸乙醇酸磷酸酯酶;③乙醇酸氧化酶;④谷氨酸乙醛酸转氨酶;⑤丝氨酸乙醛酸转氨酶;⑥甘氨酸脱羧酶;⑦丝氨酸羟甲基转移酶;⑧羟基丙酮酸还原酶;⑨甘油酸激酶 (引自李合生等,2002)
3.4.3 光呼吸的生理功能
关于光呼吸的生理功能迄今尚未未搞清楚,不同的研究者提出不同的观点。目前有两种比较流行的观点:其一是Lorimer和Andrews提出的观点,认为Rubisco同时具有羧化和加氧的功能,在有氧条件下,便不可避免地会发生加氧反应而生成乙醇酸,导致有机C的损失。C2循环的功用,是尽量回收C(75%)以避免过多的损失。这种观点的一个间接的证据是缺失C2循环的酶的突变体在光呼吸条件下不能存活,据解释是由于不能回收Rubisco加氧反应所损失的C,使叶绿体内的C耗竭,光合作用也就停止。另一个比较流行和比较多人赞同的观点是光呼吸消耗了多余能量,对光合器官起保护作用,避免引致伤害而产生光抑制。植物在强光下,如果CO2供应不足(如在干旱、盐碱等逆境加强光照时,气孔关闭使CO2不能进入),叶绿体吸收的过多的能量会对光系统Ⅱ产生伤害,从而导致光抑制。Osmond 和Bjorkman提出,由于光呼吸是一个耗能过程,消耗了多余能量,便可以避免发生光抑制。其证据是在强光下而缺CO2和O2(CO2同化和光呼吸均减弱)时,便发生光抑制。但据Orgen从化学计量学推算,以在空气中和25℃下羧化反应和加氧反应的比值为4:1计,每10分子RuBP发生羧化和加氧反应至再生成10分子RuBP共消耗了91分子ATP,而单纯羧化反应则消耗90分子ATP。认为在空气条件下进行光呼吸并没有消耗太多能量,只是同化CO2减少了。而且光抑制是在CO2浓度低于CO2补偿点下发生的。在无CO2和低O2浓度下,也能有效地防止光抑制,这也很难用光呼吸的保护作用来解释。因此,关于光呼吸的保护功能,仍有待于进一步证明。越来越多的结果证明:叶黄素循环、超氧物歧化酶及各种抗氧化剂在光保护中起作用。
不管光呼吸的生理功能如何,一个不可否认的事实是:对C3植物来说,光呼吸是一个必需的生理过程,因为光呼吸缺陷的突变体在正常空气中是不能存活的,只有在高CO2浓度下(抑制光呼吸)才能存活,说明在正常空气中光呼吸是不可缺少的。
3.4.4 光呼吸与其他代谢途径的联系
光呼吸不是一个孤立的生理过程,它和其他一些代谢过程有一定关系。
在光呼吸C2环中包括氮循环。氮循环不仅在维持C2循环的运转以回收C骨架中起重要作用,在叶子的氮代谢中也有重要作用。从光呼吸释放出CO2的量来推算,叶子中有相当大部分的氮经过氮循环。有结果表明,抑制光呼吸使豆科植物的固氮作用大大加强。这除了由于增加光合产物供根系固氮作用之外,氮循环的抑制也促进根系氮的固定。在氮循环中生成的甘氨酸和丝氨酸是蛋白质合成的原料。此外,也有相当多的试验证明,C2循环中生成的NADH,可穿梭至细胞质供硝酸还原之用。
由于光呼吸降低了光合作用,使叶子光合作用合成蔗糖减少。在严重情况下(如缺乏CO2供应或高O2浓度),甚至完全不能合成蔗糖,叶绿体内贮存的淀粉也会被动员,转变为C3植物的中间产物,最后被光呼吸氧化为CO2并释放出体外。在这种情况下,植物不能生长最后死亡。
在线粒体内,同时存在光呼吸的甘氨酸-丝氨酸转化和暗呼吸的TCA循环和电子传递链,二者之间有一定的关系。一方面,由甘氨酸氧化生成的NADH可以进入电子传递链而被氧化并生成ATP;另一方面,甘氨酸氧化需要NAD+参与,这便会与TCA环中需NAD+的脱氢酶(如异柠檬酸脱氢酶、α-酮戊二酸脱氢酶和苹果酸脱氢酶等)竞争NAD+。试验证明,在有基氨酸存在时,体离线粒体中异柠檬酸和苹果酸的氧化均受抑制。所以光呼吸对暗呼吸TCA环的运转有一定影响。
叶绿体进行光合作用需要磷的参与。光合产物以磷酸丙酮糖形式输出叶绿体外,这便需要从叶绿体外输入无机磷来补充。据报道,当用甘露醇处理以造成菠菜叶片缺磷,并从21%O2转入2%O2以抑制光呼吸,光合吸收CO2出现振荡现象。缺磷的烟草在抑制光呼吸时亦出现光合振荡。从C2循环的化学计量学来看,在整个反应过程中有Pi释放出,可以回补叶绿体内的Pi。
3.4.5 C3植物、C4植物以及CAM植物的光合特点
人们根据光合作用碳素同化的最初光合产物的不同,把高等植物分成两类:(1)C3植物。这类植物的最初产物是3-磷酸甘油酸(三碳化合物),这种反应途径称C3途径,如水稻、小麦、棉花、大豆等大多数植物。(2)C4植物。这类植物以草酰乙酸(四碳化合物)为最初产物,所以称这种途径为C4途径,如甘蔗、玉米、高粱等。一般来说,C4植物比C3植物具有较强的光合作用,其原因有结构和生理两方面。
C4植物叶片的维管束鞘薄壁细胞较大,其中含有许多较大的叶绿体,叶绿体没有基粒或基粒发育不良;维管束鞘的外侧紧密排列一层环状或近于环状的叶肉细胞,组成了“花环型”(Kranz type)结构。这种结构是C4植物的特征。叶肉细胞内的叶绿体数目较少,个体小,有基粒(图3-29)。
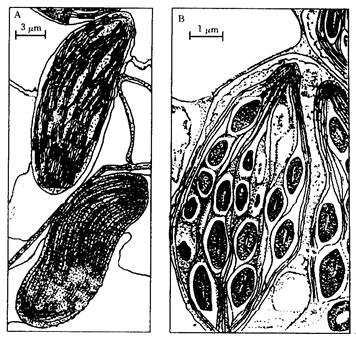
图3-29 玉米叶的两种叶绿体
A.上边是叶肉细胞内的叶绿体,有基粒;下边是维管束鞘薄壁细胞内的叶绿体,无基粒。固定前24h暗处理,所以没有淀粉粒B. 维管束鞘薄壁细胞内的叶绿体,无基粒,有淀粉粒
维管束鞘薄壁细胞与其邻近的叶肉细胞之间有大量的胞间连丝相连。C3植物的维管束鞘薄壁细胞较小,不含或很少叶绿体,没有“花环型”结构,维管束鞘周围的叶肉细胞排列松散(图3-30)。由于C4植物卡尔文循环仅在维管束鞘薄壁细胞中进行。所以,只在维管束鞘薄壁细胞形成淀粉,在叶肉细胞里没有淀粉。而水稻等C3植物由于仅有叶肉细胞含有叶绿体,整个光合过程都是在叶肉细胞里进行,淀粉亦只是积累在叶肉细胞中,维管束鞘薄壁细胞不积存淀粉。
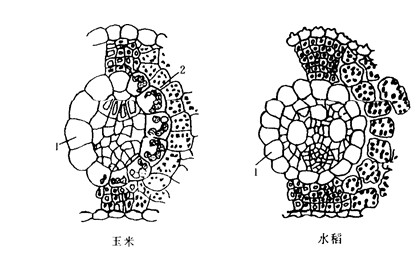
图3-30 C4植物(玉米)与C3植物(水稻)叶片解剖结构的差异
1.维管束鞘 2. 维管束鞘薄壁细胞中的叶绿体
在生理上,C4植物一般比C3植物具有较强的光合作用,这与C4植物的PEPC活性较强,光呼吸很弱有关。
前面已经提过,卡尔文循环的CO2固定是通过Rubosco来实现的,C4途径的CO2固定是由PEPC催化完成的。这两种酶都可以固定CO2形成有机物。但它们对CO2的亲和力却差异很大。PEPC对CO2的Km值(米氏常数)是7μmol,而Rubisco的Km值是450μmol。前者比后者对CO2的亲和力高很多。试验证明,C4植物PEPC的活性比C3植物高60倍。由于C4植物的PEPC活性较强,对CO2亲和力很大,加之C4二羧酸是由叶肉细胞进入维管束鞘细胞。所以,PEPC就起一个“二氧化碳泵”的作用。因此,C4植物的光合速率比C3植物高许多,尤其是在二氧化碳浓度低的环境下,相差更是悬殊。
由于PEPC对CO2的亲和力大。所以,C4植物能够利用低浓度的二氧化碳,而C3植物不能。由于这个原因,C4植物的CO2补偿点(0~10mg/L CO2)比C3植物的CO2补偿点(50~150mg/L CO2)低很多。所以,C4植物亦称为低补偿植物,C3植物亦称为高补偿植物。
由于C4植物能利用低浓度的CO2,当外界干旱气孔关闭时,C4植物就能利用细胞间隙里的含量低的CO2,继续生长,C3植物就没有这种本领。所以,在干旱环境中,C4植物生长比C3植物好。
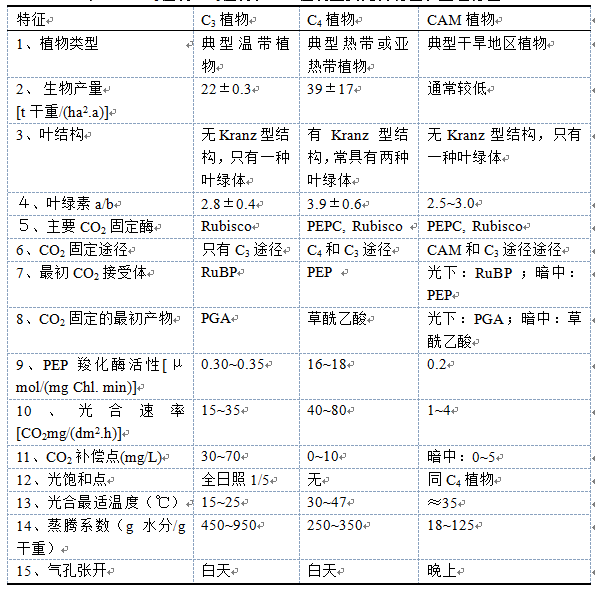
表3-5 C3植物、C4植物和CAM植物主要光合特征和生理特征
由于PEPC起“二氧化碳泵”的作用,把外界CO2 “压”进维管束鞘薄壁细胞中去,增加维管束鞘薄壁细胞的CO2/O2比率,改变Rubisco的作用方向。因为Rubisco在不同的CO2/O2情况下,产生不同的反应,具双重性。在CO2/O2比值高时,这种酶主要使核酮糖二磷酸进行羧化反应,起羧化酶作用,形成磷酸甘油酸,所以乙醇酸积累就少;在CO2/O2比值低时,这种酶主要使核酮糖二磷配进行氧化反应,起加氧酶作用,形成磷酸乙醇酸和磷酸甘油酸,产生较多的乙醇酸。由于C4植物具有“二氧化碳泵”的特点,因此,C4植物在光照下只产生少量的乙醇酸,光呼吸速率非常之低。此外,C4植物的光呼吸酶系主要集中在维管束鞘薄壁细胞中,光呼吸就局限在维管束鞘内进行。在它外面的叶肉细胞,具有对CO2亲和力很高的PEPC。所以,即使光呼吸在维管束鞘放出CO2,也很快被叶肉细胞再次吸收利用,不易“漏出”,也是C4植物的CO2补偿点低的原因之一。
综上所述,把C4植物、C3植物和CAM植物主要的光合和生理特征总结于表3-5。

