
课件:Chapter VII plant photomorphogenensis.ppt(下载附件 5.25 MB)
中文课件:
提要 光作为环境信号作于植物,调节植物生长、分化和发育过程称为光形态建成。感受光的受体有光敏色素、隐花色素和紫外光-B受体。光敏色素普遍分布于除真菌以外的低等和高等植物中,其中以分生组织中含量较多。光敏色素有两种类型:Pr型和Pfr型,二者的吸收光谱不同,并且在不同光谱作用下可以相互转换。Pr型没有生理活性,Pfr型具有生理活性。Pfr与X组分形成Pfr·X复合物后,经过信号传导途径,最终才产生生理功能。光敏色素介导广泛的生理作用, 最终影响植物的光形态建成。隐花色素和紫外光—B受体的化学本质尚不清楚,但它们对植物的生长发育都有影响。
光是影响植物生长发育的外界环境(如光、温度、重力、水分和矿物质等)中最为重要的条件。它对植物的影响主要有两个方面:第一,光是绿色植物光合作用所必需的,光合作用是高能反应,与光能的强弱有关,其光的受体是光合色素。第二,光能调节植物整个生长和发育。这种依赖光控制植物生长、发育和分化的过程,称光形态建成(photomorphogenesis)。有人也称之为光控发育。光形态建成是低能反应,所需能量比光补偿点的能量还低10个数量级。光是以信号影响植物的生长发育,与信号的有无及信号的性质(即波长)有关。植物体内接收光信号的受体是光敏色素,隐花色素和紫外光—B受体。与植物细胞中通常含有的大量光合色素相比较,这些受体的含量很微少,但是它们对外界光环境的变化很敏感。
7.1 光受体
光形态建成中光的受体是光敏色素、隐花色素和紫外光—B受体。目前对光敏色素研究较深入,下面重点介绍光敏色素。
7.1.1 光敏色素
7.1.1.1 光敏色素的发现和分布
早在上世纪二十年代就已观察到,光照与黑暗的相对长度控制着某些植物的开花。1935~1937年Flint等在研究光质对莴苣种子萌发的影响时,发现某些波长的光促进萌发,而另一些波长的光抑制萌发(图7.1)。促进莴苣种子萌发最有效的光是在红光区域(600~700nm),而抑制莴苣种子萌发的光是在远红光区域(720~740nm)。后来(1946~1960),Borthwick(植物学家)和Hendricks(物理化学家)在美国马里兰州Beltsville美国农业部实验室装置了大型光谱仪,更精确地发现促进莴苣种子萌发最有效的光是波长约为660nm的红光,抑制其萌发最有效的光是波长约730nm的远红光。他们进一步发现,如果用红光和远红光交替地照射莴苣种子,种子萌发的促进或受抑制决定于最后照射光的波长。而与交替的次数无关(表7.1)。
表7.1 交替的暴露在红光(R)和远红光(FR)下的莴苣种子萌发百分率*
| 光处理 | 萌发率 / % | 光处理 | 萌发率 / % |
| R | 70 | R-FR-R-FR | 6 |
| R-FR | 6 | R-FR-R-FR-R | 76 |
| R-FR-R | 74 | R-FR-R-FR-R-FR | 7 |
* 在26℃下,连续的以1min的R和4min的FR曝光
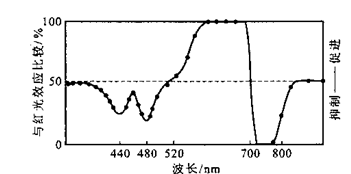
1959年同一研究小组的Butler等研制出双波长分光光度计,测定黄化芜菁子叶和黄化玉米幼苗的吸收光谱。发现幼苗经红光照射后,其红光区域吸收减少,而远红光区域的吸收增加;反之,照射远红光后,其远红光区域吸收减少,而红光区域吸收增加。红光和远红光交替照射后,这种吸收光谱可以多次可逆的变化。他们依此认为,这种吸收红光和远红光并且可以互相转化的光的受体是具有两种存在形式的单一色素。在1960年Borthwick等将这种色素命名为光敏色素(phytochrome)。其红光吸收形式为Pr(蓝绿色),远红光吸收形式为Pfr(黄绿色)。光敏色素的发现是上个世纪植物科学中的一大成就,也有人认为,光敏色素的发现是植物光生理学发展的里程碑。
目前已知除真菌外,各类植物,包括藻类、苔藓、地衣、蕨类、裸子植物和被子植物中都有光敏色素。
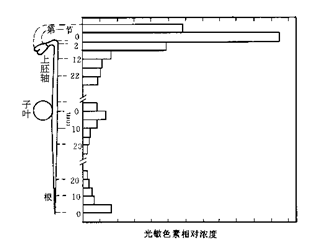
光敏色素分布在植物的各个器官中,但在植物体内的分布是不均匀的,黄化幼苗的光敏色素含量比绿色幼苗多20~100倍。禾本科植物胚芽鞘的尖端、黄化豌豆幼苗的弯钩、各种植物的分生组织具有较多光敏色素。一般来说,蛋白质丰富的分生组织中含有较多的光敏色素(图7.2)。
许多实验证明,在黑暗中生长的植物,光敏色素以Pr形式均匀地分散在细胞质中,而在照射红光后Pr即转化为Pfr并迅速地与质膜、内质网膜、质体膜、线粒体膜等结合在一起。
现已证明,光敏色素在高等植物体内至少有两种类型:一种在黄化幼苗中含量高,在光下不稳定,被称为黄化组织光敏色素(etiolated tissue phytochrome)或光不稳定光敏色素(light-labile phytochrome)或phyⅠ,它的吸收峰在666 nm;另一种在绿色组织中为主,光下相对稳定被称为绿色组织光敏色素(greentissue phytochrome)或光稳定光敏色素(light-stable phytochrome)或phyⅡ,phyⅡ的吸收峰为652nm。
7.1.1.2 光敏色素的化学性质及光化学转换
1、化学性质光敏色素是一种易溶于水的二聚体色素蛋白,两个单体中的各个单体均由生色团和脱辅基蛋白质(apoprotein)组成。
生色团(chromophore)是一个长链状的4个吡咯环,它以硫醚键结合到120~127 kDa多肽端的半胱氨酸残基上(图7.3),具有独特的吸光特性。光敏色素生色团的生物合成是在质体中黑暗条件下进行的,其合成过程可能类似脱植基叶绿素(叶绿素的前身)的合成过程。生色团在质体中合成后就被运送到细胞质中,与脱辅基蛋白质装配形成光敏色素结合蛋白(图7.4)。
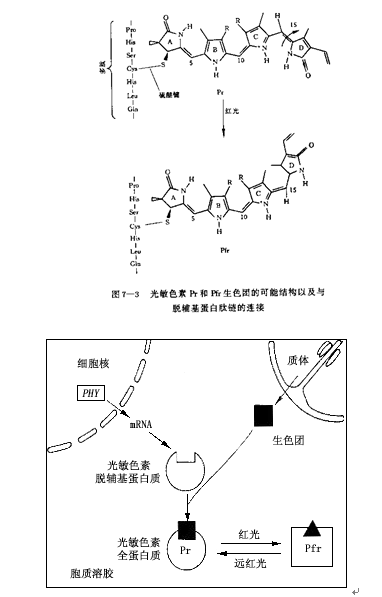
图 7.4光敏色素生色团与脱辅基蛋白质的合成与装配
2、类型 光敏色素有两种类型:红光吸收型(red light-absorbing form, Pr)和远红光吸收型(far-red light-absorbing form, Pfr)。二者光学特性不同,图7.5是光敏色素两种类型的吸收光谱,Pr吸收高峰在660nm,而Pfr的吸收高峰在730nm。二者在红光和远红光的作用下可以相互转换。当Pr吸收660nm红光后,就转变为Pfr,而Pfr吸收730nm远红光后,会逆转为Pr。Pfr是生理激活型,Pr是生理失活型。

在Pr和Pfr相互转变时,生色团和脱辅基蛋白质也发生构象变化,其中生色团变化带动脱辅基蛋白质变化,因为前者吸收光。(Pr生色团吸光后,吡咯环D的C15和C16之间的双键旋转,进行顺反异构化,这种变化导致吡咯环构象发生变化。当Pr转变为Pfr时,脱辅基Pfr也进行构象变化。)实验证明,Pr氨基末端暴露在分子表面,而Pfr的则隐蔽在内部。
近年来已在拟南芥中克隆了5个脱辅基蛋白质基因,分别为PHYA、PHYB、PHYC、PHYD和PHYE,说明高等植物光敏色素属基因家族。不同基因编码的蛋白质有各自不同的时间、空间分布,有不同的生理功能。研究表明,PHYA编码phyⅠ光敏色素,而其他4种基因编码phyⅡ光敏色素。PHYA的表达受光的负调节,在光下其mRNA的合成受到抑制。而其余4种基因表达不受光的影响,属于组成性表达。通过转基因研究发现,光敏色素蛋白质有不同的功能区域,N末端是与生色团连接的区域,与光敏色素的光化学特性有关,phyA、phyB的特异性也在此区域表现出来。C末端与信号转导有关,两个蛋白质单体的相互连接也发生在C端。
3光化学转换 Pr和Pfr在小于700nm的光波下,都有不同程度的吸收,出现相当多的重叠(图7.5)。在活体中,在一定的波长下,具有生理活性的Pfr浓度占光敏色素的总浓度(Cptot=CPr+CPfr)的比例被称为光稳定平衡(Photostationaryequilibrium ,Φ,)即Φ= CPfr /Cptot。不同波长的红光和远红光可以组合成不同的混合光,能得到各种Φ值。例如,白芥幼苗在饱和红光(600nm)下的是0.8,也就是说Pfr占总光敏色素80%,而Pr占20%;而在饱和远红光(718nm)下的Φ值是0.025,即总光敏色素的2.5%是Pfr,97.5%是Pr。在自然条件下,决定植物光反应的Φ值为0.01-0.05时就可以引起显著的生理变化。
Pr与Pfr之间的转变包括几个ms至μs的中间反应。在这些转变过程中,包括光化学反应和黑暗反应。光化学反应局限于生色团,黑暗反应只有在含水条件下才能发生。这就可以解释为什么干种子没有光敏色素反应,而用水浸泡后则有光敏色素反应。
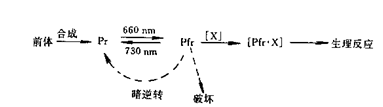
Pr比较稳定,Pfr不稳定。在黑暗条件下,Pfr会逆转为Pr,使Pfr浓度降低;Pfr也会被蛋白酶降解。Pfr一旦形成,即和某些物质(X)反应,生成Pfr·X复合物,经过一系列信号放大和转变过程,产生可观察到的生理反应。X的性质还不清楚,在具体的反应中应是信号转变链上的早期组分。从拟南芥突变体研究中发现了COP和DET基因家族,他们的基因产物是光信号传导途径的早期重要组分,位于光敏色素以及蓝光受体的下游,作为控制植物光形态发生和暗形态发生之间的开关。COP和DET突变体在黑暗条件下不表现出黄化现象。光敏色素Pr和Pfr在植物体内的相互转化及作用总结如下:
7.1.1.3光敏色素的生理作用
Pfr·X复合物,经过一系列信号放大和转变过程,引起一系列生理生化反应,最终表现为光形态建成、成花诱导等。这些生理生化反应非常广泛,包括种子萌发、幼苗的生长和发育、茎和叶的伸长、根和叶原基的分化、膜电势、叶的运动和节律现象等。当然,光敏色素通过调控基因和酶介导这些生理生化反应。
光敏色素可以调节许多酶的活性,现已发现60多种酶或蛋白质受光敏色素的调节,下面列举了一些受光敏色素调节的酶(表7.2)。
表7.2 受光敏色素调节的酶
| 叶绿体形成与光合作用: | 氨基酮戊酸脱水酶,NADPH-脱植基原叶绿素氧化还原酶,叶绿素a/b结合蛋白,铁氧还蛋白,光基因—32,3—磷酸甘油醛脱氢酶,Rubisco,PEPcase |
| 呼吸与能量代谢 | 细胞色素C氧化酶,烯醇化酶,腺苷酸激酶,二磷酸果糖激酶,延胡索酸水合酶,葡萄糖—6—磷酸脱氢酶,3—磷酸甘油醛脱氢酶,异柠檬酸脱氢酶,苹果酸脱氢酶,抗坏血酸氧化酶,过氧化氢酶,过氧化物酶 |
| 碳水化物代谢: | 淀粉酶,α—半乳糖苷酶 |
| 脂类代谢: | 脂氧合酶 |
| 氮及氨基酸代谢: | 硝酸还原酶,天冬氨酸氨基转移酶,谷氨酰胺合成酶,谷胱甘肽还原酶,苯丙氨酸转氨基酶 ,亚硝酸还原酶,谷氨合成酶 |
| 蛋白质代谢: | 氨酰t-RNA合成酶 |
| 核酸代谢 | 核糖核酸酶Ⅱ,RNA核苷酸转移酶,三磷酸核苷酶 |
| 次生代谢: | 苯基苯乙烯酮异构酶,苯基苯乙烯酮合成酶,肉桂酸羧化酶,苯丙氨酸氨基裂解酶 |
| 激素代谢与生长发育: | 吲哚乙酸氧化酶,羧甲基戊二酰—CoA还原酶,精氨酸脱羧酶 |
7.1.1.4光敏色素的作用机理
关于光敏色素的作用机理主要有两种假说:膜假说和基因调节假说。
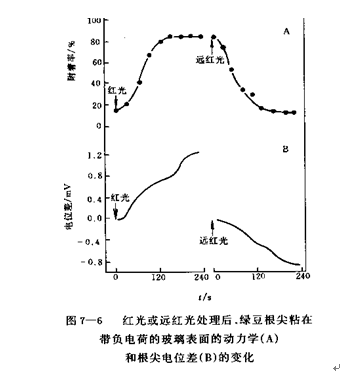
1、膜假说 膜假说由Hendricks和Borthwick在1967年提出,认为光敏色素的活跃形式Pfr直接与膜发生物理作用,通过改变膜的一种或多种特性或功能而参与光形态建成。如棚田效应(Tanada effect)。该效应表现为离体绿豆根尖在红光下诱导膜产生少量正电荷,所以能粘附在带负电荷的玻璃表面,而远红光照射则逆转这种粘附现象(图7.6)。该假说可以解释光敏色素调节的快反应。实验中发现,红光对膜电位的影响特别快,在燕麦根表面照光后4.5s就可以记录到膜的超极化,而在巨藻(Nitella)的实验中,照光后1.7s就发生去极化反应。如此快速的反应有可能是光敏色素和膜直接作用产生的。
Dreyer和Weisenseel(1979)发现,转板藻在照射30s红光后,细胞内Ca2+积累速度增加2-10倍,接着照射30s远红光,这个效应就全部逆转。由于质体和原生质膜之间存在肌动球蛋白纤维,钙调素(CaM)能活化肌球蛋白轻链的激酶在植物中也得到证明。有几个科学家提出在转板藻中从光受体到生理反应之间存在着这样一种信号链:红光→Pfr增多→跨膜Ca2+流动→细胞质中Ca2+增加→CaM活化→肌球蛋白轻链激酶活化→肌动球蛋白收缩运动→叶绿体转动。
2、基因调节假说 该假说由Mohr在1966年提出,认为光敏色素通过调节基因的表达而参与到光形态建成中。现有资料证明,光敏色素调节基因的表达都发生在转录水平上。已有试验证明,Rubisco小亚基的基因(rbcs)的表达受红光和远红光的调控,光可诱导其mRNA水平的提高。现已查明,在rbcs基因的转录起始点上游存在与光敏色素调节有关的区域,这些区域对光调节这些基因的表达很重要。
有人将光形态建成过程中基因活化概括为:光信号经过传递、放大,通过激活转录因子,活化或抑制某些特定基因,使转录出单股mRNA的速度发生改变,mRNA翻译成特殊蛋白质(酶),最后表现出形态建成(图7.7)。

7.1.2 隐花色素
上个世纪70年代以来,可见光谱中的短波光对植物生长、发育和代谢的影响日益受到重视。研究发现,蓝光和近紫外光调控着植物的许多生长发育过程,如高等植物的向光性反应,气孔的开启,叶绿体的分化和运动,茎伸长的抑制作用等专一的受蓝光、近紫外光诱导调节,而红光、远红光不起作用;除高等植物外,蕨类植物丝状体发育为原叶体、真菌菌丝体类胡萝卜素的合成及其分生孢子的分化、一些海藻配子体发生及其卵诱导产生等都需要蓝光、近紫外光的诱导。现已证实,植物界存在的第二类光形态建成反应是受蓝光、近紫外光调节,其光受体称作蓝光/近紫外光受体(B/UV-Areceptor),简称蓝光受体。Gressel提出隐花色素(Cryptochrome)一词,表示不同于光敏色素,隐花色素作用光谱的高峰处在450mm左右,所以常称隐花色素所起的作用为蓝光效应。蓝光效应一般不能被随后处理的较长波长的光照所逆转。
蓝光受体的研究近几年来虽有很大进展,但目前关于它的化学性质尚不清楚,目前倾向于认为是某种黄素蛋白。
7.1.3 紫外光——B受体
它是一类未知化学本质的光受体,它的吸收光谱处在紫外光区,即波长为280~320nm的紫外光短波区,称为紫外光——B受体(UV-Breceptor),它在细胞内吸收280~320nm紫外光引起光形态反应,它的作用高峰在290nm左右,如它诱导玉米黄化苗的胚芽鞘和高粱第一节间形成花青素,其作用光谱在290~300nm有一峰。
7.2 光形态建成
光敏色素、隐花色素等光受体,在吸收了不同波长的光以后,可以诱导和调节植物的形态建成,并对某些生理过程有着显著影响。就光敏色素而言,目前知道从种子萌发到幼苗胚芽鞘、中胚轴生长,幼叶的展开,或下胚轴伸长,弯钩伸直,两片子叶的张开和扩展;从叶片表面气孔的形成和活动到幼茎维管束的分化、表皮层的形成;从植株地上部形态建成到根的生长和根冠比调节;从叶绿体的发育到光合活性的调控;从贮藏淀粉、脂肪的降解到硝酸盐的还原同化、蛋白质、核酸的合成;从光周期控制花芽分化到叶片等器官衰老的调节,都存在着光敏色素的作用。表7.3列举了高等植物中一些由光敏色素控制的反应。
表7.3 高等植物中一些由光敏色素控制的反应
| 1.种子萌发 | 6.小叶运动 | 11.光周期 | 16.叶脱落 |
| 2.弯钩张开 | 7.膜透性 | 12.花诱导 | 17.块茎形成 |
| 3.节间延长 | 8.向光敏感性 | 13.子叶张开 | 18.性别表现 |
| 4.根原基起始 | 9.花色素形成 | 14.肉质化 | 19.单子叶植物叶片展开 |
| 5.叶分化和扩大 | 10.质体形成 | 15.偏上性 | 20.节律现象 |
现在列举不同光受体诱导和调节光形态建成的一些实例。
7.2.1 光与种子萌发
早在1907年Kinzel报道,在研究的964个植物种中,672种在光下发芽率增强,258种发芽受光抑制。近来,有人分析了142种非栽培植物,其中107个光促进发芽,3个受抑制,32个对光反应不敏感。我们把萌发时需要光的种子叫需光种子,又称喜光种子(lightfavored seeds);萌发时不需要光的种子,称为需暗种子(dark favored seeds)。近年来的进一步研究表明,种子休眠和萌发对某些波长的光较敏感,它们主要是红光、远红光和蓝光,作为光的受体一般认为光敏色素是主要的,而隐花色素对种子的休眠也有一定的调节作用。有关隐花色素的研究较少,这里主要介绍光敏色素在种子光休眠中的作用。
前面已经提到莴苣种子发芽受红光和远红光控制。表明了光敏色素参与了休眠的解除与种子的萌发。对于休眠的解除需要光的种子来说,一般均要在种子或种子特定部位处于吸胀状态时才能进行。在种子成熟后的干种子状态,其中含有光敏色素的Pr或Pfr形式,它们往往是稳定的,当这样的种子遇到适宜的萌发环境条件,在光的照射下,Pr发生水合并转换成Pfr形式,从而导致发芽;如果把莴苣种子经吸胀后,照光形成Pfr,然后立即去水合,那么这样的Pfr仍然是稳定的,在一年内再水合时仍能在暗中萌发。
实验中发现,把莴苣的胚从胚乳、种皮和果皮中分离出来,排除胚覆盖物的影响来观察照光对胚根萌发的作用。结果显示,分离的胚能在暗中生长,但用红光照射后胚根伸长快得多,当再用远红光照射时,胚根伸长减慢。这与整粒种子的情况相似。实验表明,胚根-下胚轴区可能是种子光敏感区。
在对光休眠的诱导与解除中,激素是另一个要考虑的因素。目前对于种子照光后光敏色素与几种植物激素的关系有很多报道,但缺少较一致的看法,有人认为,红光照射后形成光敏色素的活性形式Pfr,可能通过引起GA、细胞分裂素合成,或破坏ABA,从而破除光休眠的。例如用光能促使发芽的种子一般也可用GA促使萌发,GA在促进萌发上的作用与Pfr十分相似。
80年代有关GA克服光休眠的直接证据来自拟南芥突变体的研究。例如一个GA缺失的拟南芥突变体,在水中时即使照光也不发芽,但在光下用1mM GA4+7处理,或在暗中用100mM GA4+7处理时,就可以发芽。这表明GA可以克服遗传障碍,代替种子的需光要求。拟南芥菜种子萌发之所以对光有要求,可能是光能诱导一种或一种以上GA的形成。
光敏色素参与的种子光休眠与解除休眠有一定的生态学意义。一般说来,大粒种子具有足够贮藏物质以维持幼苗较长时间生长在地下黑暗环境中,它的发芽一般不需要光,如瓜类;而小粒种子,特别是一些草本植物种子当它们处于光不能透过的土层时,保持休眠状态,只有当它处于土表,依赖少量贮藏物质进行发芽,从而及时伸出土表迅速进行自养生长。反之,如果小粒种子在土表下深处黑暗中就能发芽,待它还不能伸出土表时,可能已耗尽种子贮藏物质而不能存活。
7.2.2 光与营养生长
幼苗的生长也是受光控制的。在单子叶植物,如谷类中,幼苗期开始时,胚芽鞘迅速伸长,随后伸长抑制,而幼叶从胚芽鞘中抽出,在这一系列过程中都是与光敏色素的调节有关。黄化玉米幼苗的卷叶在红光下展开,继而用远红光照射又重新卷起来。在双子叶幼苗阶段,胚轴破土伸出地表后,茎尖会形成钩状,当给以红光照射时形成的Pfr促使了钩的展开。
当幼苗生长进行光合作用,进而营养生长时,把双子叶植物置于红光下(光敏色素可起调节作用)或蓝光下(隐花色素起作用),往往抑制了茎的伸长。如果把植物移到阴影下,由于阴影下辐射的光线主要是远红光,从而使光敏色素由Pfr形式转为Pr形式,茎的伸长被大大促进。植物生长的快慢与体内生长素含量变化有关,而光照对生长素有破坏作用。光可使自由型的IAA转变为无活性的束缚型IAA,还可提高IAA氧化酶的活性,尤其是蓝紫光,其引起IAA氧化分解的作用更显著。试验指出,在IAA溶液中加入核黄素或紫黄质,则加快IAA氧化分解,因为这些色素吸收光谱的高峰在蓝紫光。光引起IAA氧化的产物主要是3-亚甲基氧代吲哚。由于蓝紫光能抑制植物的生长,这也是高山植物比平原植物生长矮的原因之一,因为高山的光照中,蓝紫光和紫外光强度比平原的光照中强。此外,低强度的光也抑制茎和禾本科中一些植物(如燕麦)的中胚轴的伸长。森林中处于林冠下生长的树木也呈现同样现象。林业生产上适当增加种植密度以获得少分枝、少结节的通直优质木材。在农业生产上,利用浅蓝色塑料薄膜低温下育秧,比无色的好,因其可大量透过400-500nm波长的蓝紫光,使秧苗矮壮;同时可吸收大量的600nm波长的橙光,使膜内温度升高,有利秧苗生长。
在一些草本植物的茎基部会出现分枝,称分蘖。分蘖的多少一定程度上取决于光敏色素的调节作用。在谷物生长十分稠密时,照到茎基部的主要是富含远红光的光,从而使Pr比例增高,分蘖形成受阻。如果用富含红光的光照射则可促进分蘖。
除上述之外,光对植物生长的许多过程如休眠芽的萌发生长,冬季植物生长减慢停止等都有影响,并且都是通过光敏色素实现的。
7.2.3 光与花色素苷和其它类黄酮物质的合成
大多数植物能在特殊的细胞中合成花青素苷和其它类黄酮物质,这个过程往往是光促进的,如苹果果实照光一侧易产生红色、秋天红叶、花瓣着色等。花青素苷产生的作用光谱显示,有效光是红光、远红光和蓝光区,而绿光几乎是无效的。在大多数植物的花色素苷合成中,光敏色素和隐花色素可能是主要光受体。
其它促进类黄酮物质合成的光的受体可能是UV-B光受体,UV-B还能诱导欧芹悬浮培养细胞大量积累黄酮类物质,研究表明黄酮生物合成的关键酶之一PAL的活性受紫外光调节。在促进类黄酮合成中,UV-B与光敏色素可能呈现协同作用。UV-B对植物细胞有一定程度的伤害作用(UV-B能降低黄瓜、向日葵、大豆、水稻等的株高、叶面积和光合能力),因此在表皮细胞中由紫外光诱导形成能够吸收UV-B的花青苷和黄酮类物质,这可能是植物一种自我保卫反应。
7.2.4 光与叶绿体的向光性反应
光可以调节叶绿体在细胞内的移动。当外界光照强度很强时,细胞中的叶绿体能在径向细胞壁上排列,并将窄面朝向阳光,使叶绿体受光面变得最小,从而防止强光伤害;反之,在弱光或黑暗时,叶绿体在横向细胞壁上排列,并将扁平面朝向阳光,以增加对光的吸收。
实验指出,在藓类和被子植物中,叶绿体向光性反应的作用光谱以蓝光区为最大,隐花色素参与了这些植物中叶绿体向光性反应,而光敏色素没有涉及。而在一些如转板藻的绿藻中,叶绿体的向光性可能与光敏色素和隐花色素均有关系,光敏色素处在转板藻细胞的质膜附近,随不同光波和方向诱导叶绿体的向光反应。
在上述植物中,叶绿体本身没有吸收引起向光性反应的光,而光在细胞的其他地方首先被吸收,引起微管和微丝相互作用,改变细胞质的流动,推动叶绿体向光性运动,使叶绿体适应外界光照条件的变化。
7.2.5 光与细胞器的形成
细胞器的形成也受到光的调节。如被子植物必须在光下才能形成叶绿体,否则只能分化不富含原片层体的黄化质体而无类囊体膜系统,同时也不能形成叶绿素,只能积累原脱植基叶绿素。只有在光下随着叶绿素的形成才能出现完善的类囊体膜系统从而发育成叶绿体。在这个转变过程中,光敏色素起了主要的作用。光敏色素还控制着膜上捕光叶绿素a/b蛋白复合体、二磷酸核酮糖羧化酶,磷酸烯醇式丙酮酸羧化酶等的形成。
种子成熟和萌发时,细胞内的线粒体也同叶绿体一样发生形态结构上的转变。随着种子的成熟,含水量逐渐降低,其膜结构和呼吸酶类消失,代谢活跃的线粒体为贮存形式的原线粒体所代替;当种子萌发并在光的触发和光敏色素控制下,才能重新转变为结构完善和代谢活跃的线粒体。
7.2.6 光与气孔开启
光是影响气孔开闭最重要的外界因子。光诱导的气孔反应依赖于保卫细胞中三种光受体:叶绿素、隐花色素和光敏色素的共同作用。用饱和的红光(250 mmol m-2s-1)照射鸭趾草叶片,维持保卫细胞叶绿体和叶肉细胞正常的光合作用,在连续的强红光背景下,30s蓝闪光(250 mmol m-2s-1)立即引起气孔导度增加,15min后达最大值。
7.2.7 光周期反应
在一天之中,白天和黑夜的相对长度称为光周期(photoperiod),光周期对植物的花诱导有着极为显著的影响(见第9章),除此之外,光周期也影响着植物其他的许多生长发育过程,如块根、块茎的形成,马铃薯块茎为短日诱导,木薯、小萝卜以及其他植物作为贮藏器官的根的形成也为短日促进,但洋葱鳞茎的形成为长日促进。 另外,许多植物的性别表现也常常受光周期的影响,光周期反应与光敏色素有着密切的关系。我国发现的光敏核不育水稻农恳58s的雄性器官发育过程,也是通过光敏色素去感受日照的长短。
关键词
光形态建成(photomorphogenesis) 光敏色素(Pr、Pfr ) 光稳定平衡 (Φ) 棚田效应(Tanada effect) 隐花色素(cryptochrome) 光受体(photoreceptors) 紫外光—B受体(UV-B receptors) 喜光种子(light favoredseeds) 需暗种子(darkfavored seeds) 脱辅基蛋白质(apoprotein) 生色团(chromophore)
思考题
1. 光受体有哪几种类型?它们参与哪些生理反应?
2. 何为光敏色素?试述其性质和作用。
3. 设计试验,证明光敏色素在植物体内的存在。
4. 试述光敏色素的特点及其在植物成花中的作用。
5. 举例说明种子的需光萌发并论述光在萌发中的作用。
6. 试述光对植物营养生长的作用。
7. 叙述光敏色素的发现过程。
8. 举例说明光敏色素的快反应和慢反应并说明作用机理。
主要参考文献
1. 潘瑞炽主编《植物生理学》(第四版) 高等教育出版社,北京2001
2. 曾广文、蒋德安主编 《植物生理学》(第一版) 中国农业科技出版社,北京2000
3. 曹仪植、宋占午主编《植物生理学》(第一版) 兰州大学出版社兰州1998
4. 余叔文、汤章城主编《植物生理与分子生物学》(第二版)科学出版社 北京1998
5. 尾田义治、刘瑞弦 译《植物光形态建成》 科学出版社 北京1981
6. 李合生主编《植物生理学》
7. Kendrick R C,Kronenberg G H. Photomorphogenesis in Plants. 2nd ed. Dordrecht: KluwerAcademic Publishers, 1994

