第五节 抗寄生虫免疫
寄生虫包括单细胞的原生动物和多细胞的蠕虫,一些是细胞外增殖,另一些在细胞内增殖。寄生虫在宿主体内生长和成孰,经常引起严重和长期的组织及器官损伤。许多寄生虫具有多种宿主,使得比仅感染人的病原体更难控制。
一、抗寄生虫免疫反应
不同的寄生虫引发不同的免疫应答类型,取决于寄生虫的大小和细胞构成以及其生活周期。通常,原生动物寄生虫趋向于诱导 Th1 应答而蠕虫感染则引起 Th2 应答。
(一)抗原生动物寄生虫免疫
1.体液免疫 与胞外菌类似,所有基干抗体的对胞外菌防御的效应机制均适用于防御小的原生动物寄生中。抗寄生中抗休介导中和作用调理吞噬并激活经典补体途径。大的原生动物可通过中性粒细胞和巨噬细胞介导的 ADCC 清除。
2.Th1
应答,巨噬细胞高度活化和IFN-γTh1应答是抗原生动物寄生虫免疫的关键,因为巨噬细胞高度活化所需IFN-γ主要来源于Th1效应细胞。像许多胞内菌一样,原生动物寄生虫被巨噬细胞吞噬后,不会在普通的吞噬体中被消化。这些寄生虫能抵抗巨噬细胞通常的呼吸爆发,只有高度活化的巨噬细胞具有足够的ROIs、RNIs、TNF,才能将这些寄生虫有效杀伤。如果高度活化的巨噬细胞不能清除感染,则会形成肉芽肿。
IFN-γ具有独特的抗原生动物效应包括对许多原生动物均有直接毒性:(2)刺激DC 和巨噬细胞产生IL-12.随之触发NK和NKT、IFN-γ的产生;③诱导感染的巨噬细胞表达iNOS,导致细胞内NO的产生.后者清除寄生中本身或感染的细胞:④上调对吞噬体的成熟重要相关酶的表达:上调被感染的巨噬细胞表面Fas的表达,可被表达FasL的T细胞杀死。需要注意的是:Th2 细胞因子(如TGF- B、IL-4、IL-10和IL-13)可抑制IFN-γ和iNOS的产生,因此Th2应答优势的个体对原生动物寄生虫感染是高度易感的。
3.CTLs和γδT细胞
如果原生动物寄生虫从巨噬细胞吞噬体逃出进人胞质,寄生虫抗原可进人内源性抗原提呈途径,成为CTL的靶穿孔素/颗粒酶介导的细胞溶解对急性原生动物感染并不高效,而是CTL分泌的IFN-γ作用最大,但穿孔素/颗粒酶介导的细胞溶解在控制原生动物感染的慢性阶段更重要。与CTL类似,活化的γsT细胞产生的IFN-γ对抗原生动物寄生虫感染具有重要作用。
(二)抗蠕虫寄生虫免疫
Th1应答是抗原生动物寄生虫免疫的关键,而Th2应答是防御大的、多细胞的蠕虫的关键。与其他类型病原体不同的是:抗蠕虫Th2
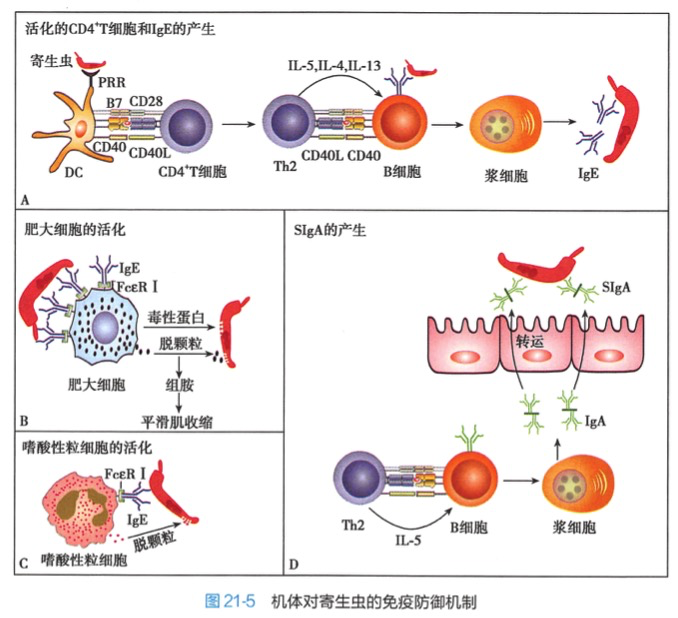
应答涉及IgE、肥大细胞和嗜酸性粒细胞。①T细胞分化为Th2细胞,后者通过CD40L-CD40相互接触使B细胞类型转换为IgE。②IgE抗体进入循环,通过结合细胞表面FcεRI“武装”肥大细胞,当蠕虫抗原结合到细胞表面的IgE时,触发肥大细胞脱颗粒,颗粒中组胺等引起宿主肠道和支气管平滑肌收缩,将寄生虫从黏膜表面驱离出宿主;此外,肥大细胞合成的组胺和其他蛋白也对蠕虫有直接毒性。③与肥大细胞脱颗粒相似,循环中的IgF可同时结合病原体和嗜酸性粒细胞表面FcεR,触发嗜酸性粒细胞脱颗粒、释放杀伤蠕虫的物质。
Th2细胞因子IL-4、IL-5和IL-13对防御蠕虫很关键。IL-4是驱动B细胞向IgE转换的主要因子;IL-5强力促进嗜酸性粒细胞的增殖、分化和活化,并支持浆细胞向IgA分化,分泌型的IgA(SIgA)抵御寄生虫进一步的黏膜定植;IL-4和IL-13抑制巨噬细胞IL-12的产生、IFN-的产生和Th1
极化;IL-13 对支气管和胃肠对寄生虫的驱离反应是必需的。

二、寄生虫的免疫逃逸机制
具有多阶段生活周期的寄生虫通过多种机制逃避免疫攻击。
1. 逃避抗体攻击
(1)抗原变换:宿主刚刚产生了针对生活周期前一阶段寄生虫表位的体液免疫应答,寄生虫发育到下一阶段,防御滞后接踵而来。如布鲁斯锥虫在某一时点仅表达其上百种VSG 基因的一种,该病原可有规则地关闭其上个VSG基因、活化另一基因,导致一种变换的球蛋白外壳,使得针对上个VSG蛋白的抗体不能识别它:其他寄生虫可通过脱落部分外膜躲避抗体的攻击。
(2)自我隔离:硕大利什曼原虫通过将自己隔离在宿主巨噬细胞中以逃避抗体攻击。
(3)伪装:血吸虫通过获得宿主糖脂和球蛋白外壳伪装自己。这种由宿主分子形成的密集“外衣”阻止抗体与寄生虫表面抗原的结合。
(4)消化抗体:一些蠕虫通过产生某种物质来消化抗体。
2.逃避吞噬溶酶体
许多原生动物发展了逃避吞噬溶酶体的方法。如:一些肠内的原生动物溶解粒细胞和巨噬细胞,使在第一现场被吞噬的机会最小化;鼠弓形体阻止巨噬细胞吞噬体融合到溶酶体;锥虫溶酶体融合之前酶解吞噬体膜,然后逃避到宿主细胞的胞浆中;硕大利什曼原虫则经常保留在吞噬体中,干预呼吸爆发。
3.逃避补体攻击原生动物和蠕虫均可通过蛋白水解的方式消除吸附到其表面的补体活化蛋白或前切寄生电结合抗体的Fc部分:也可分泌一些分子强迫液相补体活化.以耗竭补体成分;还可表达模仿哺乳动物 RCA 蛋白、DAF的蛋白以保护自身不被补体攻击。
4.干预T细胞攻击
原生动物和蠕虫均可通过干扰宿主T细胞应答来保护寄生虫的存活。例如:恶性疟原虫可促使Th细胞分泌IL-10而不是IFN-γ,导致MHCⅡ类分子的表达下调,抑制NO的产生。硕大利什曼原虫表达可结合巨噬细胞上CR3和FcvRs的分子、降低这些细胞产生IL-12、抑制
Thl 应答。钩虫分泌数种可诱导宿主T细胞低应答甚至耐受的蛋白,这种免疫抑制状态使大量的钩虫集聚在感染的宿主体内。其他一些丝虫类的蠕虫诱导
APC 下调其表面MHCI和Ⅱ类分子及其他抗原提呈基因,使这些APC 不能启动T细胞活化。

