第三节 抗胞内菌免疫
胞内菌通过损伤的皮肤黏膜或媒介的叮咬进人宿主体内后,在宿主细胞内繁殖,以逃避吞噬细胞、补体及抗体的攻击。胞内菌常见的靶细胞有上皮细胞、内皮细胞、肝细胞和巨噬细胞等。因为巨噬细胞具有运动能力,所以细菌感染巨噬细胞后可以迅速播散至全身。胞内菌在宿主细胞内繁殖,但通常毒性不强,不会产生损伤性的细菌毒素,从而与宿主细胞“共存”。其胞内生活方式使得难以从宿主体内彻底被清除,导致慢性疾病,如结核分枝杆菌导致的肺结核。
一、抗胞内菌的免疫
抗胞内菌免疫机制与抗胞外菌的最大不同是:抗胞外菌主要依赖体液免疫,而抗胞内菌主要依赖细胞免疫。
(一)抗胞内菌固有免疫
1.中性粒细胞和巨噬细胞的作用 最早到达感染局部的是中性粒细胞。中性粒细胞分泌防御素破坏尚未进入宿主细胞的胞内菌,从而控制早期感染;有些细菌虽然逃脱了防御素的破坏,随之可被中性粒细胞吞噬后通过强大的呼吸爆发杀灭。活化的巨噬细胞在吞噬及杀灭胞内菌的过程中起着重要作用。巨噬细胞还通过TLR对分枝杆菌的脂蛋白和脂多糖识别,活化产生促炎细胞因子,促进NK细胞活化和Th1细胞分化,进而杀灭细菌。
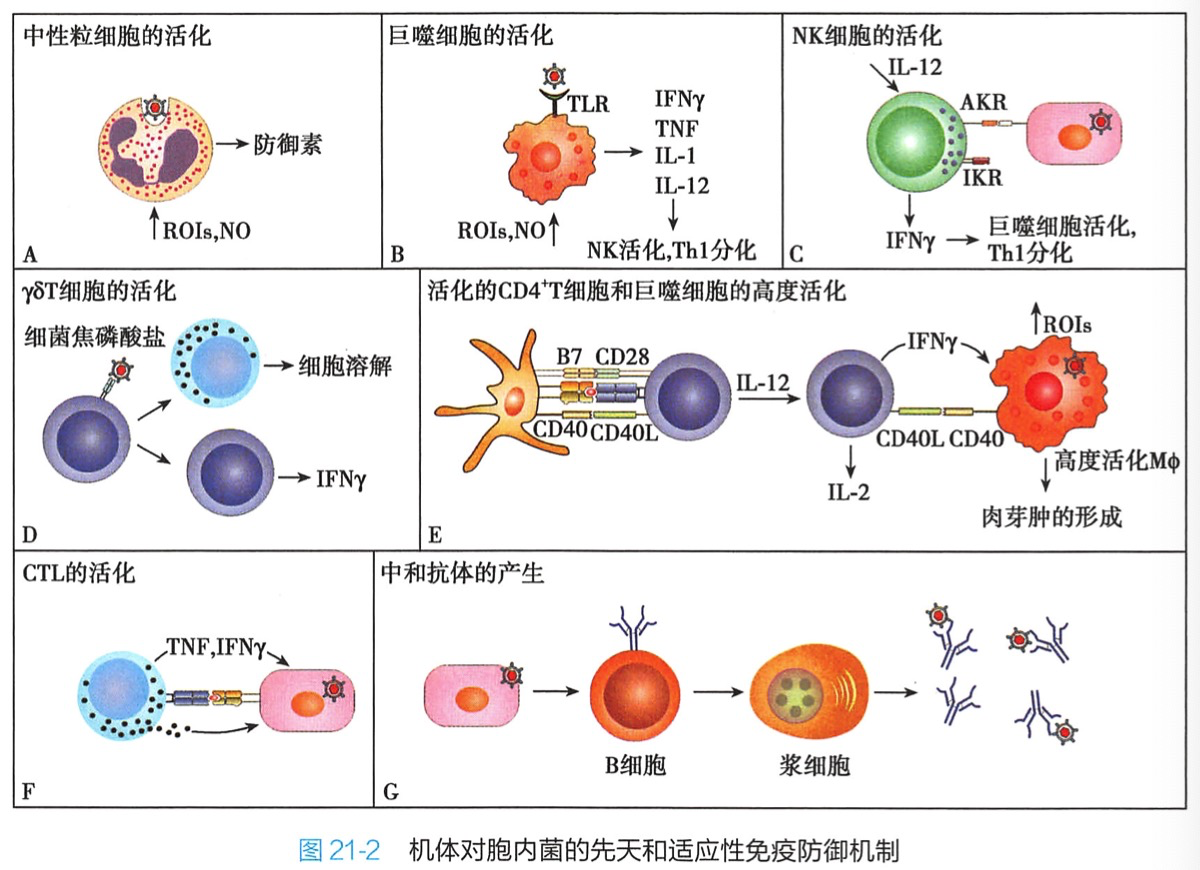
2.NK细胞和γδT细胞的作用 在巨噬细胞等协同下,NK细胞被活化杀伤宿主细胞。活化的NK细胞分泌大量的IFN-γ,促进巨噬细胞活化、间接促进Th1细胞分化。γδT细胞会识别胞内菌如分枝杆菌的小磷酸化分子,引发效应γδT细胞增殖,通过杀伤或分泌IFN发挥抗菌效应。
(二)抗胞内菌适应性免疫
1.CD8+T细胞应答 CTL 细胞对清除胞内菌感染起关键作用。DC获取了由被吞噬细菌降解或宿主细胞死亡而产生的抗原,通过抗原交叉提呈激活 CTL。细菌蛋白通过内源性抗原提呈途径成为CTL细胞的靶标。胞内菌特异性CTL很少通过Fas介导的细胞凋亡途径或穿孔素介导的细胞溶破作用杀伤靶细胞,而主要通过分泌 TNF、IFN-γ和(或)具有直接杀菌活性的颗粒清除靶细胞。
2.CD4+T细胞应答 胞内菌活化的特异性CD4+T细胞效应为:分化为Th1细胞释放IFN-γ辅助巨噬细胞活化,后者产生大量ROI和RNI,发挥强大的抗菌作用。
抗胞内菌应答中Th1应答比Th2 应答更重要。如在麻风患者中,Th2应答上调的患者易患破坏性的麻风病,即瘤型麻风;而 Th1 应答上调的患者麻风病症状减轻,即结核样麻风。
3.抗体应答 细菌特异性中和抗体,虽然不能直接清除胞内菌,但可与尚未进入细胞的细菌结合,或与释放到胞外环境中但还没有感染新的宿主细胞的子代菌结合,阻断细菌进入宿主细胞,并通过调理吞噬或补体介导的溶菌作用清除胞内菌。
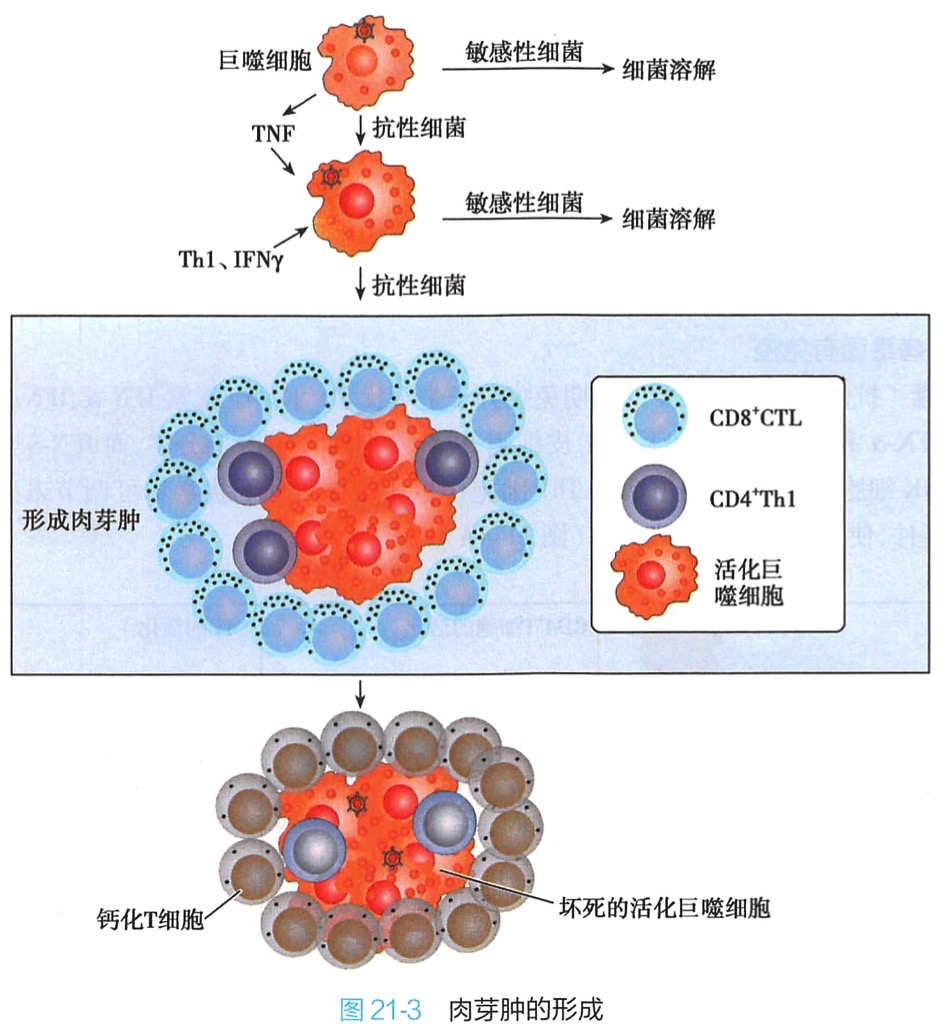
(三)肉芽肿的形成
当宿主抗胞内菌免疫与病原体的博弈相持不下、而转为慢性感染时,就会在宿主感染局部形成一种称为肉芽肿的结构以局限化感染。肉芽肿的内层包含巨噬细胞和CD4+T细胞,而外层是CD8+T细胞。部分肉芽肿的外层钙化、纤维化、中间的细胞坏死,死亡细胞中的所有病原体都被杀灭,感染被消除;少数病原体仍然存活并在肉芽肿中休眠,如果肉芽肿破裂,病原体就会被释放,重新开始增殖。如果宿主的免疫应答处于免疫抑制状态,无法聚集抵抗新一次攻击所必需的T细胞和巨噬细胞,病原体可能进入血液,进一步感染全身的组织,甚至导致死亡。
二、胞内菌的免疫逃逸机制
像胞外菌及其他病原体一样,在宿主免疫压力下,胞内菌也进化出逃避免疫的机制。胞内菌多为慢性感染,其逃避免疫的能力更强、机制更为复杂。
1.逃避吞噬杀伤 某些胞内菌可选择在非吞噬细胞中增殖,以逃避吞噬杀伤,例如麻风分枝杆菌会感染人体外周神经的施万细胞。另有一些胞内菌可使吞噬细胞失活,或逃避吞噬细胞的杀伤,如李斯特杆菌进入吞噬细胞后合成李斯特杆菌溶血素 O(LLO),破坏吞噬溶酶体,使细菌逃逸到胞质中。
2.逃避抗体的中和作用 一些胞内菌通过细胞-细胞接触机制进入另一个宿主细胞,使中和抗体无法发挥中和作用。如:李斯特杆菌可诱导宿主产生基于肌动蛋白的伪足,内陷进入邻近的非吞噬细胞,由此细菌不会暴露到胞外,逃避抗体中和作用。
3.阻止淋巴细胞活化 某些胞内菌通过干预APC的抗原提呈功能、阻止淋巴细胞活化而逃避T细胞杀伤。如:结核分枝杆菌感染 DC后会引起 MHCI类分子、Ⅱ类分子和 CD1的下调,使抗原无法有效提呈和活化T细胞、NKT细胞。


