第二节 免疫学发展简史
免疫学的发展经历了长期的过程,从早期对免疫学的朦胧认识到目前对免疫学比较系统的认识, 该发展过程是连续和渐进的。 我们人为地将此过程划分为三个时期,即经验免疫学时期、 实验免疫学 时期和科学免疫学时期。
一、经验免疫学时期
人类对免疫的认识首先是从与传染病作斗争中开始的。 人类观察到传染病患者在痊愈之后可以 抵抗该种传染病再次侵袭,我国古代医学家将此现象称为“以毒攻毒”,由此开始尝试通过人工轻度 感染某种传染病以获得对该种传染病的抵抗力。 例如,葛洪所著的《肘后备急方》 (约公元303 年)和 孙思邈所著的《备急于金要方》 (约公元648 年)对于防治狂犬病就有"取狂犬脑敷之,后不复发”的文 字记载,可以说,是我国古代医学家在国际上第一次进行了“预防接种"的实践。 天花曾是一种烈性 传染病,因其通过呼吸道传播,人是唯一的易感宿主,死亡率极高,严重威胁人类的生存。 例如,18 世纪发生在欧洲的天花大流行,就造成6000 万人死亡。 据考证,公元16 世纪我国明朝隆庆年间已有关 于种痘的医书记载,将天花患者康复后的皮肤痘皮磨碎成粉,吹入未患病的儿童的鼻腔可预防天花。
这种种痘的方法不仅在当时国内广泛应用,还传到俄国、 朝鲜、日本、 土耳其和英国等国家。 早在1772 年英国王室就开始允许在英国小孩中采用此法。 据记载,天花流行时,种过痘的人群死亡率只有不接 种人群的1/10 到1/5。 种人痉预防天花具有一定的危险性,但为日后牛疮苗的发现提供了宝贵的经 验(图 1-1) 。

公元18 世纪后叶,英国医生Edward Jenner 观察到挤牛奶女工接触患有牛痘的牛后,可被传染却不会再得天花。 他意识到人工接种“牛痘”可能会预防天花,于是,用两年时间在24 名志愿者身上进 行了接种“牛痘"预防天花的试验,取得了成功。 1798 年Jenner 出版了相关专著,提出了"vaccination" 的概念(vacca 在拉丁语中是牛的意思,意为接种牛痘),开创了人工主动免疫的先河。 人类经过将近 180 年的努力,于1980 年世界卫生组织(WHO)庄严宣布,全球已经消灭了天花,这是一个具有划时代 意义的人类医学历史的伟大事件,彰显了免疫学对于人类健康的巨大贡献。
二、实验免疫学时期
(一)实验免疫学的兴起
免疫学发展的初期主要是抗感染免疫。 病原菌的发现和疫苗的研制推动了免疫学的发展。 19世纪70 年代许多致病菌陆续被分离成功,德国细菌学家Robert Koch 提出了病原菌致病的概念,颠覆 了先前入类对瘟疫"的认识。 在此基础上,进一步认识到将减毒的病原体给动物接种,可预防有毒 的病原体感染所引起的疾病。 法国微生物学家和化学家Louis Pasteur 发现炭疽杆菌经40 -43'C 较高 温度下培养后,可明显降低毒力,将其制成人工减毒的活菌苗接种牲畜可预防炭疽病的发生,其后Pasteur 又将狂犬病病原体经过兔脑连续传代获得减毒株,制备成减毒狂犬疫苗。 在随后的20 多年时 间里,随着越来越多的致病菌被确定,多种多样的疫苗(vaccine) 也相继间世。
(二)细胞免疫和体液免疫学派的形成
19 世纪后叶,俄国学者Elie Metchnikoff 发现吞噬细胞可吞噬异物,千1883 年提出了细胞免疫的假说,即吞噬细胞理论。 他高瞻远瞩地推测,吞噬细胞是天然免疫中的重要部分,其对获得性免疫也 至关重要,并提出,炎症并不是单纯的一种损伤作用,也是机体的一种保护机制。 这一理论对生物学 和医学的发展产生了深远而广泛的影响。 Metchnikoff 的伟大发现开创了固有免疫,并为细胞免疫奠 定了基础。 经过近百年的努力,人们对参与固有免疫的细胞和分子,固有免疫细胞识别外来病原生物 的机制,固有免疫应答的特点,以及固有免疫与适应性免疫的关系都有了深入的了解。
1890 年,Emil von Behring 和他的同事Kitasato Shibasaburo 将白喉外毒素给动物免疫,发现在免疫动物血清中产生了一种能中和外毒素的物质,称为抗毒素。 次年,他们用白喉抗毒素血清成功地救治了一名患白喉的儿童。 白喉抗毒素的问世,挽救了成于上万患儿,开创免疫血清疗法即人工被动免疫 的先河,也兴起了体液免疫的研究。 1901 年von Behring 成为第一届诺贝尔生理学或医学奖得主。 抗 毒素发现后不久,又相继在动物免疫血清中发现有溶菌素、凝集素、沉淀素等特异性组分,并能与相应 的细胞、微生物及其产物发生特异性结合。 其后将血清中多种不同的特异性反应物质称为抗体(anti body), 而将能诱导抗体产生的物质统称为抗原(antigen) , 建立了抗原抗体的概念,并陆续建立了体外 检测抗原或抗体的多种血清学技术。1899 年比利时医生Jules Bordet 发现在可以溶解细菌的新鲜免 疫血清中,还存在一种热不稳定的物质,在抗体存在的条件下,具有溶菌或溶细胞的作用,这种非特异 性的物质称为补体(complement ) 。
免疫化学的研究使人们在分子水平上对抗原决定簇和抗原抗体结合的特异性开始有了认识。 20 世纪初,Karl Landsteiner 把称为半抗原的芳香族有机分子偶联到蛋白质分子(载体)上,以此为抗原免 疫动物,发现抗原特异性是由抗原分子表面特定的化学基团所决定,开启了抗体与半抗原关系的研究 领域。 此后,Landsteiner 进一步发现人红细胞表面糖蛋白所连接糖链末端寡糖结构的差异决定了 ABO 血型,并将此成果应用于临床,避免了不同血型输血引起的输血反应,极大地推动了临床医学的 发展。 Landsteiner 是血型血清学的奠基者,先后发现了ABO、MNP 和Rh 等血型系统。
1937 年Ame Tiselius 和Elvin Kabat 利用蛋白电泳的方法,发现免疫血清中丫球蛋白水平显著升 高,并具有明显的抗体活性。 据此,他们认为抗体就是丫球蛋白。 事实上,丫球蛋白组分中富含抗体, 但a 和B 球蛋白中也有部分抗体。
1959 年,英国生物化学家Rodney Porter 和美国生物化学家Gerald Edelman 各自对免疫球蛋白化 学结构进行了研究,阐明了免疫球蛋白的单体是由一对轻链和一对重链借二硫键连接在一起,免疫球 蛋白分子的氨基端组成了能与抗原特异性结合的Fab 片段,不能结合抗原但易发生结晶的狻基端称 为Fe 段。 通过对免疫球蛋白分子重链和轻链氨基酸组成特点的研究,发现了可变区和恒定区,为以 后抗体多样性形成机制的研究奠定了理论基础。
(三)免疫学重大学说和理论
鉴于抗体的广泛作用,科学家们对于抗体产生的机制进行了深入研究,提出了不同学说,如1897年Paul Erhlich 提出的抗体产生的侧链学说(side ch扣n theory) , Linus Pauling 等提出的模板学说等,这 些学说从不同的侧面解读了抗体产生的机制,为后续研究提供了借鉴。
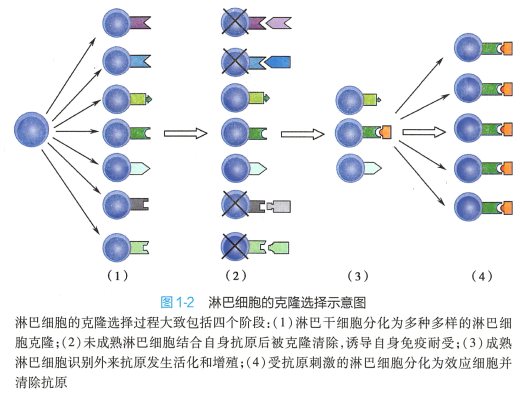
1957 年澳大利亚免疫学家Macfarlane Burnet 提出的克隆选择学说(clonal selection theory) 是免疫 学发展史中最为重要的理论。 该理论的提出,主要源千对天然免疫耐受和人工免疫耐受实验结果的 分析和思考。 1945 年Ray Owen 发现因为牛胎盘独特的组织结构,异卵双生、胎盘融合的小牛个体内, 两种血型的红细胞共存而不引起免疫反应,在体内形成了血型嵌合体。 英国免疫学家Peter Medawar 等人在1953 年应用小鼠皮片移植的实验模型,成功地进行了人工免疫耐受的实验,即新生鼠或胚胎 期如接受了另一品系的组织抗原刺激(注射脾细胞),成年后该品系小鼠移植的皮片能长期存活,而 对其他无关品系移植的皮肤仍然发生强烈的排斥反应。 Medawar 认为,动物胚胎期或新生期接触抗 原,可使其发生免疫耐受,使动物到成年期对该抗原发生特异性的不应答。 Burnet 的克隆选择学说认 为,全身的免疫细胞是由众多识别不同抗原的细胞克隆所组成,同一种克隆细胞表达相同的特异性受 体,淋巴细胞识别抗原的多样性是机体接触抗原以前就预先形成的,是生物在长期进化中获得的。 抗 原侵入后,机体只是从免疫细胞库中选择出能识别这种抗原的相应的淋巴细胞克隆,并使其活化、增 殖,扩增出许多具有相同特异性的子代细胞,产生大量特异性抗体,清除入侵的抗原。 机体自身的组 织抗原成分在胚胎期就被相应的细胞克隆所识别,这些细胞克隆产生特异性免疫耐受,赋予机体免疫 系统区分“自己”和“非己"的能力。 实际上,在胚胎期任何进入机体的抗原都将被视为自身成分而产 生免疫耐受(图1-2) 。
Burnet 克隆选择学说中提出的一个细胞克隆产生一种特异性抗体的预见,在1975 年被Georges Kohler 和Cesar Milstein 所创立的B 淋巴细胞杂交瘤技术和产生的单克隆抗体所证实。 他们设计了一种选择性培养基,只能使一种酶缺陷的骨髓瘤细胞与抗原活化B 淋巴细胞融合后形成的杂交瘤细胞 得以生长,通过克隆化方法,使一个杂交瘤细胞扩增成一个克隆(一个无性繁殖的细胞群)。 正如预 期的那样,同一个克隆的杂交瘤细胞产生抗体的特异性都是相同的。 由千单克隆抗体高度的均一性, 并能获得针对人们所需要的一种分子甚至一个抗原决定簇的抗体,加之杂交瘤具有在体内、体外无限 生长的能力,使单克隆抗体技术在生命科学和医学领域中引发了一场革命。
Niels Jerne 在1974 年提出了抗体分子上的独特型和抗独特型相互识别而形成免疫网络。 免疫网 络学说认为,抗原刺激机体产生抗体,抗体分子上的独特型决定簇在体内又能引起抗独特型抗体的产 生,抗独特型抗体又可引起针对此抗独特型抗体的抗体,如此下去,在抗体和淋巴细胞中产生一个复 杂的级联网络,在免疫应答调节中起着重要作用。 免疫网络学说丰富了免疫学理论体系。
(四)免疫学的细胞学基础的奠定
20 世纪下半叶开始,细胞免疫学开始兴起,从而对免疫系统有了全面的认识。 1957 年,Bmce Glick 发现切除鸡的富含淋巴细胞的腔上痰,导致抗体产生缺陷,遂将此类淋巴细胞称为腔上褒依赖 的淋巴细胞,简称为B 淋巴细胞或B 细胞(B 为腔上襄Bursa 的首字母)。 1961 年,Jacques Miller 采用 新生期小鼠切除胸腺的模型,Robert Good 在临床上观察一新生儿先天性胸腺缺陷,都发现了外周血 和淋巴器官中淋巴细胞数量减少,免疫功能明显缺陷,并将依赖于胸腺发育的淋巴细胞称为T 淋巴细 胞或T 细胞(T 为胸腺thymus 的首字母)。 其后不久,其他的科学家进一步证实:T 细胞负责细胞免疫 (如移植排斥),B 细胞负责体液免疫;T 细胞和B 细胞之间有协同作用,T 细胞可辅助B 细胞针对某些抗原产生IgG, 胸腺依赖抗原(即T 细胞依赖抗原)的概念也随之产生;T 细胞是一个不均一的细胞 群,有辅助T 细胞(Th) 和细胞毒T 淋巴细胞(CTL),并发现具有抑制作用T 细胞亚群(如调节性T 细 胞)的存在。
20 世纪70 年代,在肿瘤免疫研究中发现了一群预先不需抗原刺激、在无抗体存在条件下即可杀 伤肿瘤细胞的淋巴细胞,称为自然杀伤细胞(NK 细胞)。 1973 年美国学者Ralph Steinman 发现了树突 状细胞,随后的研究证实树突状细胞是功能最强的抗原提呈细胞,能够有效刺激初始T 细胞。 单核细 胞穿过内皮细胞进入组织脏器成为巨噬细胞,是同一个细胞谱系发育的不同阶段,提出了单核吞噬细 胞系统(mono-phagocytic system, MPS) , 改变了以往的网状内皮细胞系统的概念。 进一步研究发现,T 细胞中的"γδT 细胞和NKT 细胞以及B 细胞中的B-1 亚群主要参与固有免疫应答。
三、科学免疫学时期
1953 年James Dewey Watson 和Francis Cri ck 揭示了遗传信息携带者DNA 的双螺旋结构,开创了 生命科学的新纪元。 分子生物学的迅速兴起,极大地推动了免疫学的发展,不仅大量的免疫分子的基因被克隆,新的免疫分子被表达,而且使得人们对免疫应答的研究深入到基因水平和分子水平,分子 免疫学也应运而生,而且成为免疫学诸多分支中的核心。
( —) 抗体多样性和特异性的遗传学基础
1978 年日本分子生物学家Susumu Tonegawa 应用基因重排技术,揭示出免疫球蛋白C 区和V 区 基因在胚系的DNA 中是分隔的,而V 区包括了被分隔的数目众多的V 基因、D 基因和J 基因片段。 V、 D、J 基因片段的重排是产生抗体多样性的最重要的机制。 而C 基因片段则决定了免疫球蛋白的 类、亚类和型,相同的VDJ 按一定顺序分别与不同的C 基因片段的重组是免疫球蛋白类别转换的遗 传学基础。 膜型免疫球蛋白分子是B 细胞抗原识别受体。 Tonegawa 对有关免疫球蛋白基因结构和重 排的理论,对日后T 细胞受体基因结构和重排的发现产生了重要影响。
(二) T 细胞抗原受体的基因克隆
在lg 基因结构和重排发现后不久,1 984 年Mark Davis 和 Chien Saito 等成功克隆了T 细胞受体(TCR) 的基因。 TCR (3 链基因与免疫球蛋白重链基因,TCR a 链基因与免疫球蛋白轻链基因的结构 和重排有着惊入的相似,而且TCR 的多样性数目可能比BCR 还要多。 TCR 的发现为后续T 细胞杂交 瘤和T 细胞克隆技术的产生奠定了基础。
(三) MHC 限制性的发现
George Snell 在20 世纪30 年代起建立了一套同类系小鼠品系,以此为模型,发现了在同种移植排斥反应中起重要作用的基因区域,称为H-2, 继而证实了H-2 是由许多密切连锁基因组成的复合体, 每个基因座上有多个等位基因存在,这些基因称为主要组织相容性复合体(MHC) 。 MHC 是哺乳动物 基因中基因组数量最多、结构最复杂的基因群。 MHC 的基因型和表型在群体中具有高度的多态性, 正是这种多态性造成了不同个体之间识别抗原肤能力的差别,由此也决定了在群体中不同个体对同 一种抗原(如病原微生物)免疫应答能力的差别。
到了50 年代法国科学家Jean Dausset 在入体上发现了与 H-2 复合体相似的人类白细胞抗原 (HLA) 系统。 以后陆续鉴定出多种人类HLA 抗原。 Baruj Benacer叫应用不同品系的动物研究发现, 对某一特定抗原的免疫应答能力受到免疫应答基因(Ir 基因)所控制,并证明了Ir 基因位于小鼠H-2 中 1 区。 1974 年,Peter Doherty 和Rolf Zinkernagel 揭示了细胞毒性T 细胞在识别病毒感染细胞的病 毒抗原时存在MHC 限制性。 这些工作为临床肾脏移植和骨髓移植的成功奠定了重要的理论基础。
MHC 从发现到其基因结构、编码蛋白分子的结构和功能的阐明经历了半个多世纪,分子生物学 技术的应用,尤其是人类基因组计划的完成,使MHC 的遗传密码得以全面被破译。
(四 ) 细胞因子及其受体
20 世纪80 年代先后克隆出许多有重要生物学功能的细胞因子,它们在造血、细胞活化、生长和分化、免疫调节、炎症等许多重要生理和病理过程中发挥重要作用。 到了90 年代,由于人类基因组计划 的突飞猛进以及生物信息学的应用,入们对新的细胞因子及其受体结构和功能的研究,达到了前所未 有的高度,而且迅速被应用到临床医学中,成为免疫生物治疗的一项重要内容。
(五)固有免疫识别理论
1989 年Charles Janeway 提出了固有免疫的模式识别理论,1994 年Polly Matzinger 以模式识别理论为基础进一步提出了“危险模式”理论。 认为固有免疫细胞通过其表达的模式识别受体(pattern recognition receptor, PRR) , 选择性地识别病原体及其产物所共有的高度保守的分子结构(非已成分), 即病原体相关模式分子(pathogen associated molecular pattern, PAMP) 后,吞噬病原体、加工与提呈抗 原,并在危险信号的参与下启动适应性免疫应答。 该理论从新的角度解释了免疫系统为什么针对病原体入侵和组织损伤产生应答,而不对正常自身组织产生应答(即保持免疫耐受)。
(六)免疫细胞受体信号转导的研究
免疫细胞通过其膜表面的免疫受体(如TCR、 BCR、 NK 受体、固有免疫模式识别受体、细胞因子受 体、黏附分子以及死亡受体等),感应来自细胞外或细胞内的各种刺激。 这些刺激参与或调节免疫应 答必须与上述相应受体结合后,通过受体介导的信号途径,调节特定基因的表达。 免疫细胞的信号转 导途径十分复杂,不同免疫膜分子介导的信号途径各不相同,反映免疫应答和免疫调节的复杂性。 而 且不同信号途径之间存在着交互作用(cross-talking) , 在信号转导水平上形成了网络。 免疫细胞信号 转导途径的下游是通过活化特定的转录因子,使其进入胞核,调控基因的表达。 值得注意的是,不同 的信号途径可以激活相同的转录因子,可谓是"殊途同归"。 进入21 世纪之后,固有免疫受体介导的 免疫细胞活化及其信号转导机制的研究是生物医学领域的一个前沿热点,2011 年度诺贝尔生理学或 医学奖颁发给了这个领域的免疫学家。

