T细胞的表面分子及其作用
T细胞表面具有许多重要的膜分子,它们参与T细胞识别抗原,活化、增殖、分化,以及效应功能的发挥。其中,一些膜分子还是区分T细胞及T细胞亚群的重要标志。
一、TCR-CD3 复合物
1.TCR的结构和功能T细胞通过TCR 识别抗原。与BCR 不同,TCR并不能直接识别抗原表面的表位,只能特异性识别APC或靶细胞表面提呈的抗原肽-MHC分子复合物(pMHC)。因此,TCR识别pMHC 时具有双重特异性,即既要识别抗原肽,也要识别自身 MHC 分子的多态性部分,称为MHC 限制性(MHC restriction)。
TCR的每条肽链的胞膜外区各含1个可变(V)区和1个恒定(C)区。V区中含有3个互补决定区(CDR1、CDR2和CDR3)是TCR识别pMHC的功能区。两条肽链的跨膜区具有带正电荷的氨基酸残基(赖氨酸或精氨酸)通过盐桥与CD3分子的跨膜区连接.形成TCR-CD3复合体(图10-4)。构成TCR的两条肽链的胞质区很短,不具备转导活化信号的功能。TCR识别抗原所产生的活化信号由 CD3 转导至T细胞内。
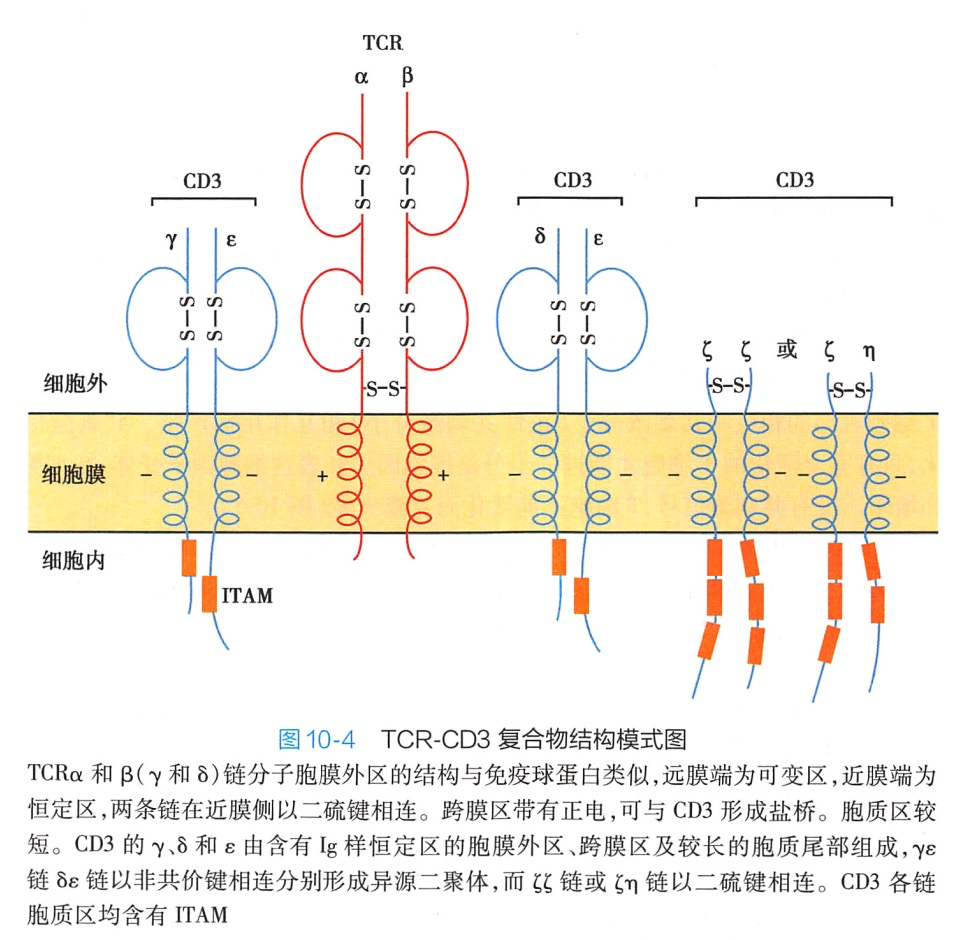
2.CD3的结构和功能 CD3具有五种肽链,即γ、δ、ε、ζ和n链,均为跨膜蛋白,跨膜区具有带负电荷的氨基酸残基(天冬氨酸),与TCR跨膜区带有正电荷的氨基酸残基形成盐桥(图10-4)。y、8和ε链的胞膜外区各有一个Ig样结构域。通过这些结构域之间的相互作用,分别形成ve和δs二聚体。ζ和ŋ链的胞膜外区很短,并以二硫键连接,形成ζζ二聚体或ζn 二聚体。γδεζ和n肽链的胞质区均含有免疫受体酪氨酸活化基序(immunoreceptor tyrosine-based activation motif,ITAM)。ITAM由18个氨基酸残基组成,其中含有2个YxxL/V(即酪氨酸-2个任意氨基酸-亮氨酸或缬氨酸)保守序列。该保守序列的酪氨酸残基(Y)被细胞内的酪氨酸蛋白激酶磷酸化后,可募集其他含有SH2结构域的酪氨酸蛋白激酶(例如ZAP-70),通过一系列信号转导过程激活T细胞。ITAM的磷酸化和与 ZAP-70的结合是T细胞活化信号转导过程早期阶段的重要生化反应之一。因此,CD3分子的功能是转导TCR识别抗原所产生的活化信号。
二、CD4 和 CD8
成熟T细胞只表达CD4或CD8,即CD4*T细胞或CD8*T细胞。CD4和CD8的主要功能是辅助 TCR识别抗原和参与T细胞活化信号的转导,因此又称为TCR的共受体。
CD4是单链跨膜蛋白,胞膜外区具有4个Ig样结构域,其中远膜端的2个结构域能够与MHCI类分子B2结构域结合。CD8是由a和β肽链组成的异二聚体.2条肽链均为跨膜蛋白,由二硫键连接,膜外区各含1个Ig样结构域,能够与 MHCI类分子重链的a3 结构域结合。
CD4和CD8分别与MHCⅡ类和MHCI类分子的结合,可增强T细胞与APC或靶细胞之间的相互作用并辅助TCR 识别抗原。CD4和CD8的胞质区可结合酪氨酸蛋白激酶 p56*。p56*激活后,可催化CD3 胞质区ITAM中酪氨酸残基的磷酸化,参与TCR识别抗原所产生的活化信号的转导过程。 CD4还是人类免疫缺陷病毒(HIV)的受体。HIV的gp120蛋白结合CD4是HIV侵人并感染CD4*T细胞或 CD4*巨噬细胞的重要机制。
三、共刺激分子
共刺激分子(co-stimulatorymolecule)是为T(或B)细胞完全活化提供共刺激信号的细胞表面分子及其配体。根据功能可将其分为正性共刺激分子和负性共刺激分子(也称共抑制分子);根据分子结构可将其分为免疫球蛋白超家族(IgSF)肿瘤坏死因子超家族(TNFSF)和整合素家族。初始T细胞的完全活化需要两种活化信号的协同作用。第一信号(或称抗原刺激信号)由TCR识别APC提呈的pMHC 而产生经CD3转导信号CD4或CD8起辅助作用第一信号使T细胞初步活化,代表适应性免疫应答严格的特异性。第二信号(或称共刺激信号)则由APC或靶细胞表面的共刺激分子与T细胞表面的相应的共刺激分子(正性共刺激分子)相互作用而产生。共刺激信号使T细胞完全活化,只有完全活化的T细胞才能进一步分泌细胞因子和表达细胞因子受体,在细胞因子的作用下分化和增殖。没有共刺激信号,T细胞不能活化而克隆失能(图10-5)。
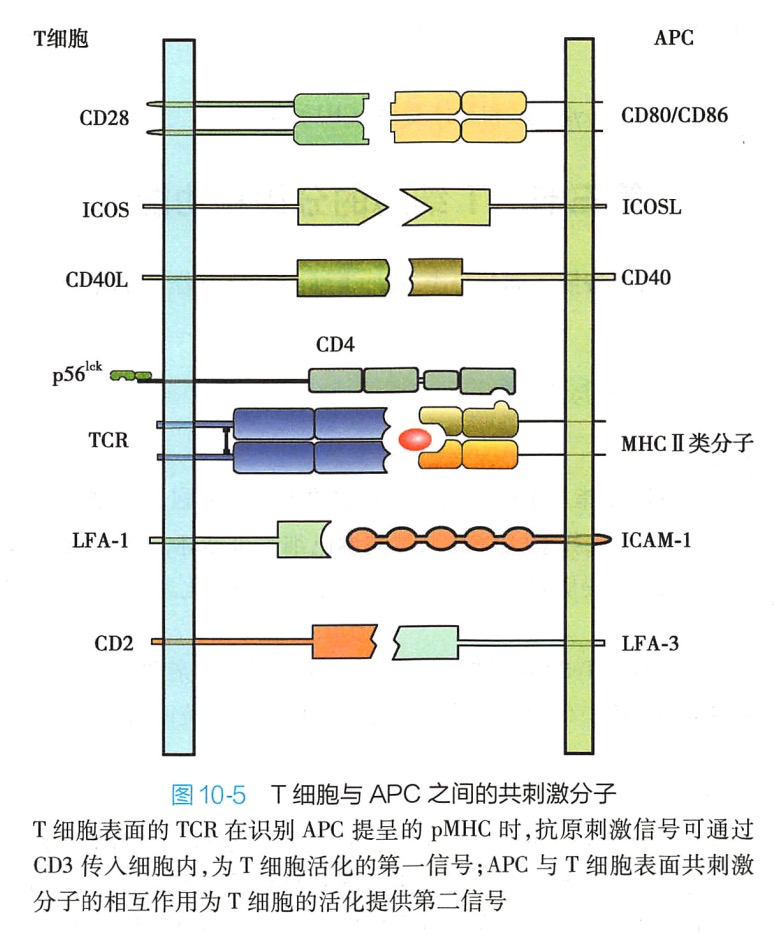
T细胞表面的正性共刺激分子主要包括:CD28家族成员(CD28和ICOS)、CD2和ICAM等,其分子结构均属于IgSF;CD28家族的配体为CD8O(B7-1)、CD86(B7-2)、ICOSL、PD-L1和 PD-L2 等。此外,正性共刺激分子还有 CD40L、FasL 以及LFA-1等,根据分子结构,前两者属于TNFSF成员,后者属于整合素家族成员。
除了正性共刺激分子外,有些表面分子可以提供免疫抑制信号,称为负性共刺激分子或共抑制分子。T细胞表面的共抑制分子主要有CTLA-4和PD-1等,根据分子结构二者均属于IgSF成员;其配体分别为 CD80、CD86 和 PD-L1、PD-L2。
1.CD28 是由两条相同肽链组成的同源二聚体,表达于90%CD4T细胞和50%CD8T细胞。CD28 的配体是CD80和 CD86.后者主要表达干专职性 APC。CD28产生的共刺激信号在T细胞活化中发挥重要作用:诱导T细胞表达抗细胞凋亡蛋白(Bcl-XL等),防止细胞凋亡;刺激T细胞合成IL-2等细胞因子,促进T细胞的增殖和分化。
2.CTLA-4(CD152)CTLA-4是重要的共抑制分子,表达于活化的 CD4 和CD8T细胞,其配体亦是CD80和CD86,但 CTLA-4 与配体结合的亲和力显著高于 CD28。由于CTLA-4的胞质区有免疫受体酪氨酸抑制基序(immunoreceptortyrosine-based inhibitory motif,ITIM),故传递抑制性信号。通常T细胞活化并发挥效应后才表达 CTLA-4,所以其作用是下调或终止T细胞活化。
3.ICOS(inducibleco-stimulator)表达于活化T细胞,配体为ICOSL。初始T细胞的活化主要依赖CD28提供共刺激信号,而ICOS则在CD28之后起作用调节活化T细胞多种细胞因子的产生,并促进T细胞增殖。
4.PD-1 PD-1(programmed death1)是重要的共抑制分子.表达干活化T细胞.配体为PD-L1和PD-L2.PD-1与配体结合后,可抑制T细胞的增殖以及IL2和IEN-v等细胞因子的产生.并抑制B细胞的增殖、分化和Ig 的分泌。PD-1 还参与外周免疫耐受的形成。
5.CD2又称淋巴细胞功能相关抗原2(LFA-2),配体为LFA-3(CD58)或CD48(小鼠和大鼠)。CD2表达于95%成熟T细胞、50%~70%胸腺细胞以及部分NK细胞.除介导T细胞与APC或靶细胞之间的黏附外,还为T细胞提供活化信号。
6.CD40 配体CD40配体(CD40LCD154)主要表达于活化的CD4*T细胞,而CD40表达于 APC。CD40L与CD40的结合所产生的效应是双向性的。一方面,促进APC活化,促进CD80/CD86表达和细胞因子(例如IL-12)分泌。另一方面,也促进T细胞的活化。在TD-Ag诱导的免疫应答中,活化Th细胞表达的CD4OL与B细胞表面的CD40的结合可促进B细胞的增殖、分化、抗体生成和抗体类别转换,诱导记忆B细胞的产生。
7.LFA-1和ICAM-1T细胞表面的淋巴细胞功能相关抗原-1(LFA-1)与APC表面的细胞间黏附分子-1(ICAM-1)相互结合,介导T细胞与APC或靶细胞的黏附。T细胞也可表达ICAM-1同APC、靶细胞或其他T细胞表达的LFA-1结合。
四、丝裂原受体及其他表面分子
T细胞还表达多种丝裂原(mitogen)受体,丝裂原可非特异性直接诱导静息T细胞活化和增殖(见第三章)。T细胞活化后还表达多种与效应功能有关的分子,例如,与其活化、增殖和分化密切相关的细胞因子受体(IL-1R、IL-2R、IL-4R、IL-6R、IL-7R、IL-12R、IFN-yR和趋化因子受体等)及可诱导细胞凋亡的 FasL(CD95L)等。
T细胞也表达 Fc 受体(如FcvR 等)和补体受体(CR1)等。

