T淋巴细胞(Tlymphocyte)来源于胸腺(Thymus),故称T细胞。成熟T细胞定居于外周免疫器官的胸腺依赖区,它们不但介导适应性细胞免疫应答,在胸腺依赖性抗原诱导的体液免疫应答中亦发挥重要的辅助作用,所以T细胞在适应性免疫应答中占据核心地位。T细胞缺陷既影响机体细胞免疫应答,也影响体液免疫应答,可导致对多种病原微生物甚至条件致病微生物(如白色念珠菌和卡氏肺囊虫)的易感性、抗肿瘤效应减弱等病理现象。
第一节 T细胞的分化发育
骨髓多能造血干细胞(hematopoietic stem cellHSC)在骨髓中分化成淋巴样祖细胞(lymphoid progenitor cell)。淋巴样祖细胞可经血液循环进入胸腺,在胸腺中完成T细胞的发育,成为成熟T细胞,再随而循环进人外周淋巴器官,主要定居干外周淋巴器官的胸腺依赖区 接受抗原制激发生免疫应答。整个过程中T细胞在胸腺中的发育至关重要。
一、T细胞在胸腺中的发育
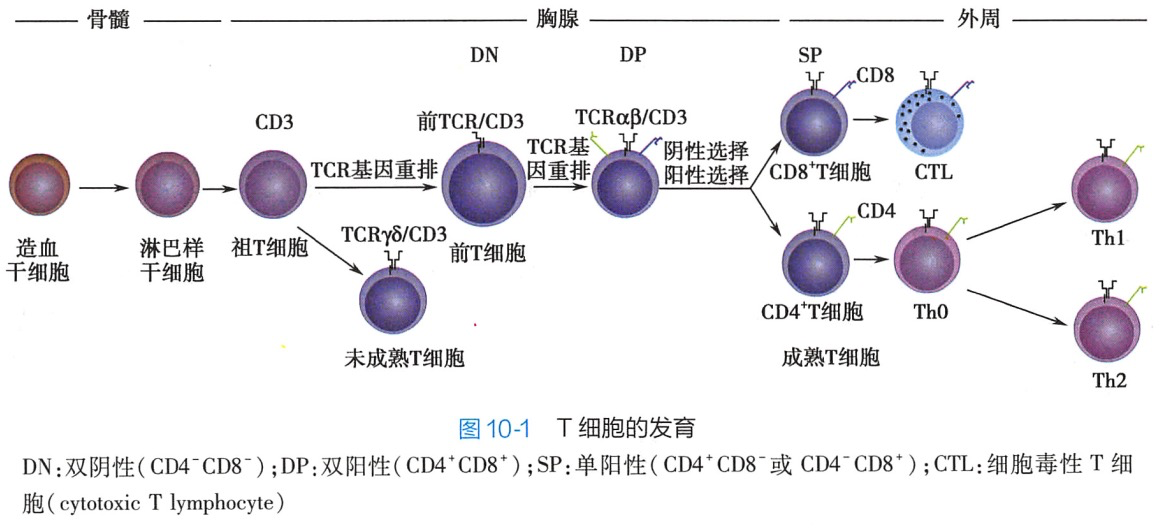
正常机体的成熟T细胞既要对多样性的非己抗原发生免疫应答,又要对自身抗原发生免疫耐受。为达到此要求,在胸腺T细胞的发育过程中,首先要经历其抗原识别受体(TCR)的基因重排,表达多样性的TCR.然后经历阳性选择和阴性选择。TCR是由α、β肽链或γ、δ肽链构成的异二聚体。T细胞在胸腺中发育的最核心事件是获得多样性 TCR 的表达、自身MHC 限制性(阳性选择)以及自身免疫耐受(阴性选择)的形成(图10-1)。
(一)T细胞在胸腺中的发育和 TCR的重排
在胸腺微环境的影响下,T细胞的发育经历淋巴样祖细胞→祖T细胞(pro-Tcell)→前T细胞(pre-T)→未成熟T细胞→成熟T细胞等阶段,不同阶段T细胞表达不同的表型并具有不同的功能。依据CD4和CD8的表达.胸腺中的T细胞又可分为双阴性细胞(double negative cell DN细胞)、双阳性细胞(doublepositive cell.DP细胞)和单阳性细胞(single positivecell,SP细胞)三个阶段。
CD4-CD8-双阴性细胞阶段 pre-T以前的T细胞均为DN细胞。其中pro-T开始重排TCR基因。根据TCR的组成.T细胞可分为表达αβTCR的T细胞和表达γδTCR的T细胞,分别简称αβT细胞和γδT细胞。γδT细胞重排γ和δ链基因;而αβT细胞重排α和β链基因,此处是γδT细胞和αβT细胞分化的分支点。在胸腺中,αβT细胞约占T细胞总数的95%~99%,γδT细胞约占1% ~5%。αβT细胞表达的β链与前T细胞α链(pre-T cella,pTα)组装成前TCR(pTα:β),成功表达前 TCR的细胞即是pre-T。在IL-7等细胞因子的诱导下,pre-T增殖活跃,并表达CD4和CD8,细胞进人 DP 细胞阶段。
2.CD4+CD8+双阳性细胞阶段DP的pre-T细胞停止增殖,开始重排α基因,并与β链组装成 TCR(a:βTCR)。成功表达TCR的细胞即是未成熟T细胞。未成熟T细胞经历阳性选择并进一步分化为 SP 细胞。
3.CD4+CD8-或CD4-CD8+单阳性细胞阶段 SP细胞经历阴性选择后成为成熟T细胞,通过血液循环进人外周免疫器官。
(二)T细胞发育过程中的 αβTCR 基因重排
TCR基因群与BCR基因群的结构相似,其重排的过程也相似。TCRβ基因群包括 Vβ、Dβ和Jβ三类基因片段。重排时先从Dβ和Jβ中各选1个片段,重排成D-J,然后与Vβ中的1个片段重排成 V-D-J.再与Cβ重排成完整的β链,最后与pTa组装成前TCR,表达于pre-T表面。TCRa基因群包括 Vα 和Jα两类基因片段。重排时从Vα和Jα中各选1个片段,重排成 V-J,再与Cα重排成完整的α链最后与β链组装成完整的TCR表达于未成孰T细胞表面(图10-2)。TCR的多样性形成机制主要是组合多样性和连接多样性,但其N序列插人的概率远高于BCR和Ig,故TCR的多样性可达10^16而此阶段的 BCR 多样性只有 10^11。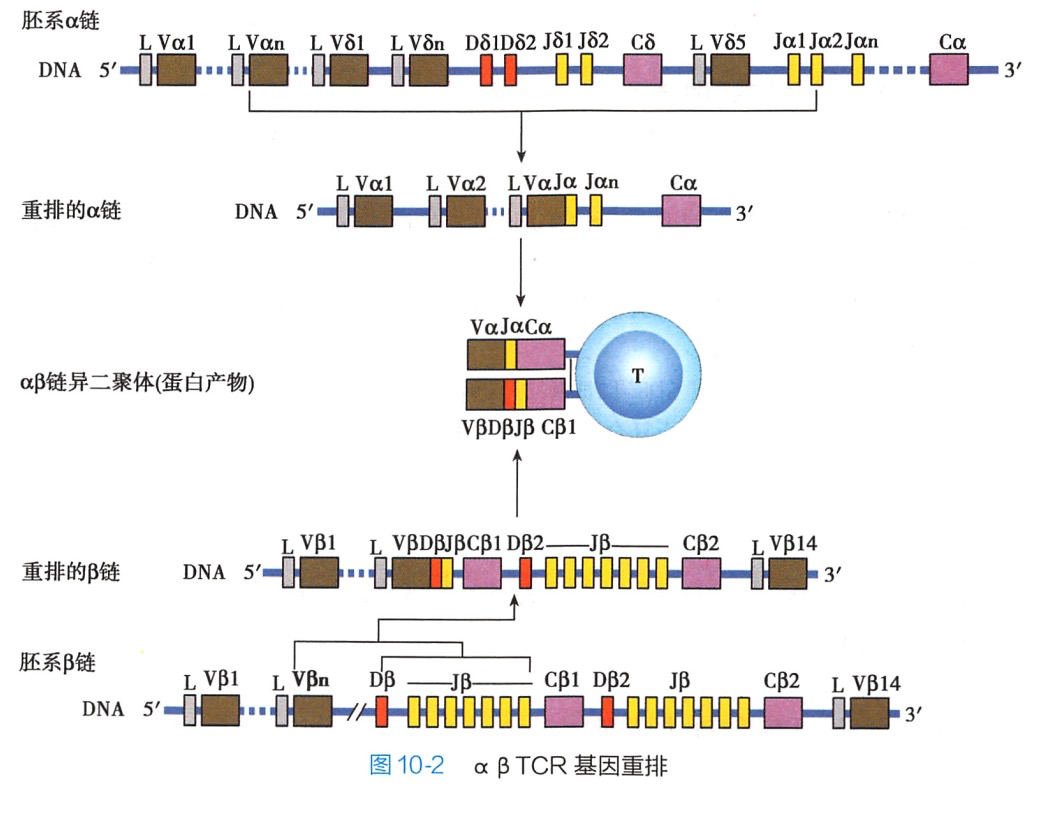
(三)T细胞发育过程中的阳性选择
阳性选择(positive selection)指在胸腺皮质中,未成熟DP细胞表达的随机多样特异性的TCR与胸腺上皮细胞表面的自身抗原肽-自身 MHCI类分子复合物或自身抗原肽-自身 MHCⅡ类分子复合物相互作用,能以适当亲和力结合(阳性)的 DP 细胞成活并获得 MHC限制性:不能结合或结合亲和力过高的DP细胞发生凋亡,凋亡细胞占DP细胞的95%以上。在此过程中DP细胞分化为SP细胞。与类分子结合的DP细胞 CD8 表达水平升高.CD4表达水平下降直至丢失:而与Ⅱ类分子结合的DP 细胞CD4表达水平升高,CD8表达水平下降最后丢失。因此,阳性选择的意义是.①获得MHC 限制性;②DP细胞分化为SP 细胞(图10-3)。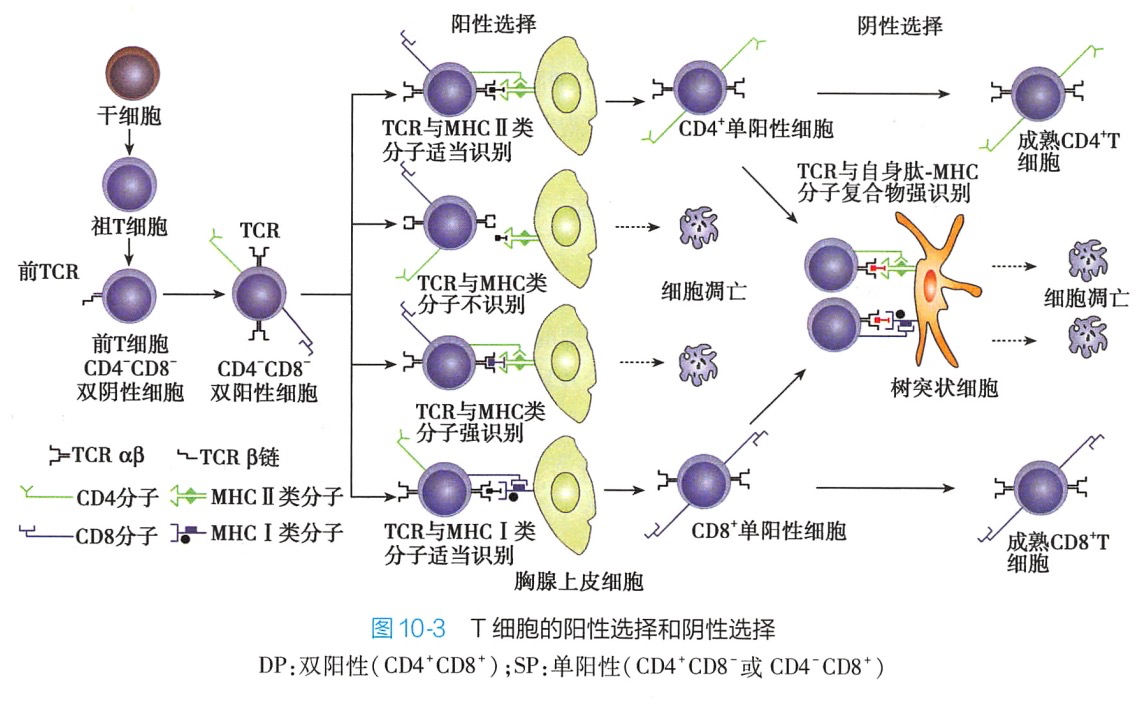
(四)T细胞发育过程中的阴性选择
阴性选择(negative selection)经过阳性选择的SP细胞在皮质髓质交界处及髓质区与胸腺树突状细胞、巨噬细胞等表面的自身抗原肽-MHCI类分子复合物或自身抗原肽-MHCⅡ类分子复合物相互作用,高亲和力结合的SP细胞(即自身反应性T细胞)发生凋亡,少部分分化为调节性T细胞;而不能结合的SP细胞(阴性)存活成为成熟T细胞并进入外周免疫器官。因此,阴性选择的意义是清除自身反应性T细胞,保留多样性的抗原反应性T细胞,以维持T细胞的中枢免疫耐受。
经过胸腺发育的CD4T细胞或CD8T细胞,进入胸腺髓质区,成为能特异性识别抗原肽-MHCI类分子复合物或抗原肽-MHCI类分子复合物、具有自身 MHC限制性以及自身免疫耐受性的初始T细胞,迁出胸腺,进入外周淋巴组织(图10-3)。
二、T细胞在外周免疫器官中的增殖分化
从胸腺进入外周免疫器官尚未接触抗原的成熟T细胞称初始T细胞,主要定居干外周免疫器官的胸腺依赖区。T细胞的定居与它在胸腺发育中获得相应的淋巴细胞归巢受体(如L-选择素等黏附分子和 CCR7等趋化因子受体)有关。T细胞在外周免疫器官与抗原接触后,最终分化为具有不同功能的效应T细胞、调节性T细胞或记忆T细胞。

