B细胞的表面分子及其作用
B细胞表面有众多的膜分子,它们在B细胞识别抗原、活化、增殖,以及抗体产生等过程中发挥作用。
一、B细胞抗原受体复合物
B细胞表面最重要的分子是 BCR 复合物。BCR复合物由识别和结合抗原的 mIg 和传递抗原刺激信号的 Iga/Igβ(CD79a/CD79b)异二聚体组成(图9-5)。
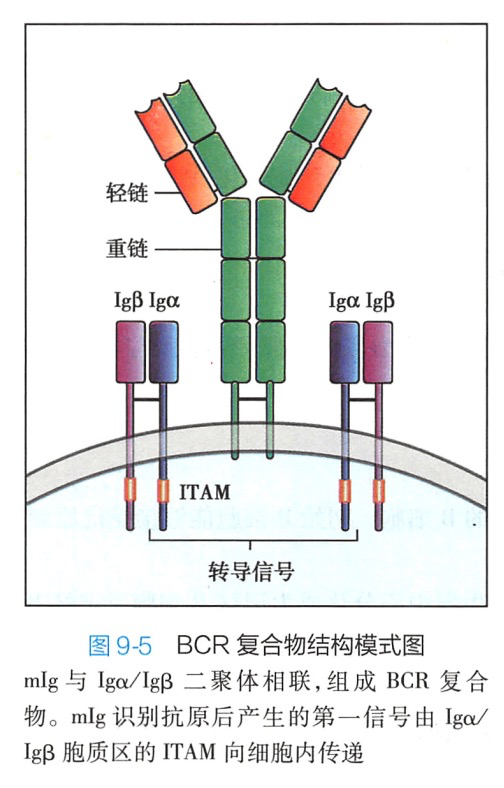
膜表面免疫球蛋白 mIg是B细胞的特征性表面标志。mIg 以单体形式存在,能特异性结合抗原,但由于其胞质区很短,不能直接将抗原刺激的信号传递到B细胞内,需要其他分子的辅助来完成 BCR 结合抗原后信号的传递。在抗原刺激下,B细胞最终分化为浆细胞,浆细胞不表达 mIg。
2.lgα /lgβ(CD79a/CD79b)Igα和Igβ均属于免疫球蛋白超家族,有胞外区、跨膜区和相对较长的胞质区。Igα 和 IgB 在胞外区的近胞膜处借二硫键相连,构成二聚体。Igα/Igβ和 mIg 的跨膜区均有极性氨基酸,借静电吸引而组成稳定的 BCR 复合物。Iga/IgB 胞质区含有免疫受体酪氨酸激活基序(immunoreceptor tyrosine- based activation motif,ITAM),通过募集下游信号分子,转导抗原与 BCR 结合所产生的信号。二、B 细胞共受体
B细胞(co-receptor)共受体能促进 BCR对抗原的识别及B细胞的活化。B细胞表面的 CD19与CD21 及 CD81 非共价相联,形成B细胞的多分子共受体,能增强BCR与抗原结合的稳定性并与Iga/Igβ共同传递B细胞活化的第一信号。在复合体中,CD21(即 CR2)可结合C3d,形成CD21-C3d-抗原-BCR复合物,发挥B细胞共受体的作用;CD19传递活化信号。此外,CD21 也是 EB 病毒受体,与 EB 病毒选择性感染B细胞有关。
三、共刺激分子
抗原与B细胞的BCR结合.所产生的信号经由Iga/IgB和CD19 转导至细胞内。此即为B细胞活化的第一信号,但仅有第一信号不足以使B细胞活化,还需要第二信号(共刺激信号)。第二信号主要由Th细胞和B细胞表面的共刺激分子(co-stimulatorymolecule)间的相互作用产生。在共刺激信号的作用下,B细胞活化增殖产生适应性体液免疫应答。而作为APC.B 细胞可以通过共刺激分子促进T细胞的增殖。
1.CD40 CD40属肿瘤坏死因子受体超家族(TNFRSF),组成性地表达于成熟B细胞。CD40的配体(CD40L即CD154)表达于活化T细胞。CD40与CD40L的结合是B细胞活化的最重要的第二信号,对B细胞分化成熟和抗体产生起重要的作用。
2.CD80和CD86CD80(B7-1)和CD86(B7-2)在静息B细胞不表达或低表达,在活化B细胞表达增强,它与T细胞表面的CD28和CTLA-4相互作用,CD28提供T细胞活化的最重要的第二信号,CTLA-4 抑制T细胞活化信号。
3.黏附分子 Th细胞对B细胞的辅助以及活化B细胞向T细胞提呈抗原均需要细胞间的接触,黏附分子在此过程中起重要的作用。表达于B细胞的黏附分子有ICAM1(CD54)、LFA-(CD11a/CD18)等,这些黏附分子也具有共刺激作用。四、其他表面分子
1.CD19 BCR识别抗原中关键的信号传递分子,也是B细胞表面特异性标志,可作为免疫治疗B细胞白血病的靶点。
2.CD20表达于除浆细胞外的各发育阶段的B细胞,可调节钙离子跨膜流动,从而调控B细胞的增殖和分化,是B细胞淋巴瘤治疗性单抗识别的靶分子。
3.CD22 特异性表达于B细胞,其胞内段含有ITIM,是B细胞的抑制性受体,能负调节 CD19/ CD21/CD81共受体。
4.CD32 有a、b两个亚型,其中CD32b即FcyRIIB,能负反馈调节B细胞活化及抗体的分泌。

