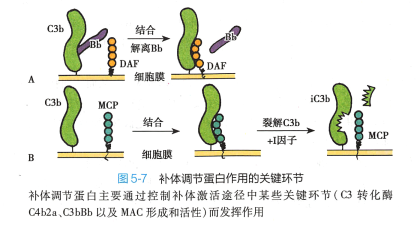
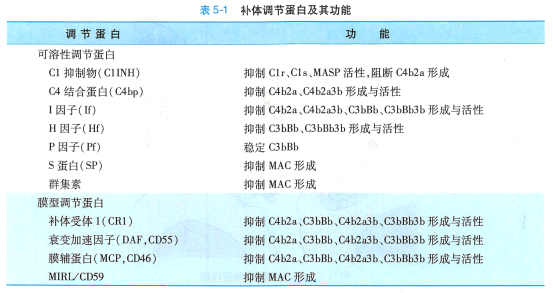
机体对补体系统活化存在着精细的调控机制,主要包括: 1)控制补体活化的启动;2)补体活性片 段发生自发性衰变;3)血浆和细胞膜表面存在多种补体调节蛋白,通过控制级联酶促反应过程中酶活 性和MAC 组装等关键步骤而发挥调节作用(图5-7 ) 。 各类补体调节蛋白针对补体激活途径关键环节 的调节机制如表5-1 。
(一)针对经典途径前端反应的调节机制
C4b2a 是经典途径和凝集素途径的C3 转化酶。 针对C4b2a 的调节因子均发挥负调控作用,主要是阻断C4b2a 形成,或分解已形成的C4b2a, 使之灭活。 另外,C5转化酶C4b2a3b也受此机制调控。 在该环节起作用的补体调节蛋白有C1抑制物(C1 inhibitor,ClINH) 、 CR1 、 C4 结合蛋白(C4 binding protein , C4 hp) 、膜辅蛋白(membrane co-factor protein, MCP) 、 I因子、衰变加速因子(decay-accelerating factor, DAF)等。
(二)针对旁路途径前端反应的调节机制
多种调节蛋白可调控旁路途径C3 转化酶(C3bBb) 形成,或抑制已形成C3 转化酶的活性。 旁路途径C5转化酶C3bBb3b 也受此机制调控。 此外,P 因子起正调节作用。
(三)针对MAC的调节机制
补体活化的共同末端通路中,多种补体调节蛋白可抑制MAC 形成和活性,从而保护自身正常细胞免遭补体攻击。 这些因子包括膜反应性溶破抑制物(membrane inhibitor of reactive lysis, MIRL) 、同 源限制因子(homologous restriction factor, HRF)亦称C8 结合蛋白(CS-binding protein, C8bp) 、 S 蛋白(S protein , SP) 、 群集素(clusterin) 等。
此外,病原体能产生一些物质抑制补体活化,逃避补体系统的攻击。 如脑膜炎奈瑟氏菌产生的H因子结合蛋白(fHbp) 及其外膜蛋白PorA 可分别将H 因子和C4bp 募集到细菌表面,灭活黏附其表面 的C3b;金黄色葡萄球菌分泌的补体抑制因子SCIN(staphylococcal complement inh加tor) 能结合C4b2a、 C3bBb, 抑制C3 转化酶活化等。

