抗体的功能
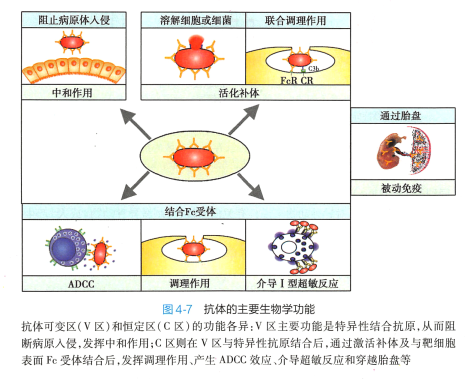
抗体的功能与其结构密切相关。 抗体分子的V 区和C 区的氨基酸组成及顺序的不同,决定了它们功能上的差异;许多不同的抗体分子在V 区和C 区结构变化的规律性,又使得抗体的V 区和C 区 在功能上有各自的共性。 V 区和C 区的作用,构成了抗体的生物学功能(图4-7 ) 。
—、抗体V 区的功能
识别并特异性结合抗原是抗体分子的主要功能,执行该功能的结构是抗体V 区,其中 CDR 在识 别和结合特异性抗原中起决定性作用。 抗体分子有单体、二聚体和五聚体,因此结合抗原表位的数目 也不相同。 Ab 结合抗原表位的个数称为抗原结合价。 单体Ab 可结合2 个抗原表位,为双价;分泌型 lgA 为4 价;五聚体lgM 理论上为10 价,但由于立体构型的空间位阻,一般只能结合5 个抗原表位,故为5 价。
抗体的V 区在体内可结合病原微生物及其产物,具有中和毒素、阻断病原入侵等免疫防御功能, 但抗体本身并不能清除病原微生物。 B 细胞膜表面的IgM 和IgD 等lg 构成B 细胞的抗原识别受体 (B cell receptor, BCR) , 能特异性识别抗原分子。 在体外可发生各种抗原抗体结合反应,有利于抗原 或抗体的检测和功能的判断。
二、抗体C 区的功能
(一)激活补体
抗体与相应抗原结合后,可因构型改变而使其C11 2 和C总结构域内的补体结合位点暴露,从而通过经典途径激活补体系统,产生多种补体的效应功能。 其中IgM 、 IgGl 和 IgG3 激活补体的能力较强, IgG2 较弱。 IgA、lgE 和IgG4 本身难以激活补体,但形成聚合物后可通过旁路途径激活补体系统。
(二)结合Fe 受体
IgG、 IgA 和IgE 抗体可通过其Fc 段与表面具有相应Fc 受体(FcR) 的细胞结合,产生不同的生物学作用。 IgG、lgA 和lgE 的Fc 受体分别称为FcγR, FcαR 和FcεR。
调理作用 (opsonization) 细菌特异性的lgG( 特别是lgGl 和IgG3) 以其Fab 段与相应细菌的抗原表位结合,以其Fc 段与巨噬细胞或中性粒细胞表面的Fc-yR 结合,通过IgG 的"桥联”作用,促进吞噬细胞对细菌的吞噬(图4-7) 。
2. 抗体依赖的细胞介导的细胞毒作用 (antibody-dependent cell-mediated cytotoxicity,ADCC) 抗体的Fab 段结合病毒感染的细胞或肿瘤细胞表面的抗原表位,其Fc 段与杀伤细胞(NK 细胞、巨噬细胞等)表面的FcR 结合,介导杀伤细胞直接杀伤靶细胞(图4-7) 。 NK 细胞是介导ADCC 的主要细胞(见第十四章) 。 抗体与靶细胞上的抗原结合是特异性的,而表达FcR 细胞的杀伤作用是 非特异性的。
3. 介导I 型超敏反应 lgE 为亲细胞抗体,可通过其Fc 段与肥大细胞和嗜碱性粒细胞表面的高 亲和力lgE Fc 受体(FcεR I)结合,并使其致敏。 若相同变应原再次进入机体与致敏靶细胞表面特异 性lgE 结合,即可促使这些细胞合成和释放生物活性物质,引起I 型超敏反应(见第十八章)。
(三)穿过胎盘和黏膜
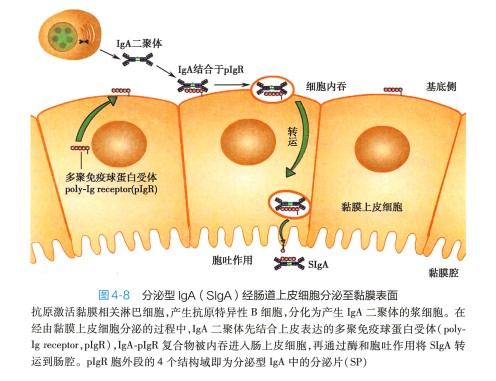
在人类,lgG 是唯一能通过胎盘的免疫球蛋白。 胎盘母体一侧的滋养层细胞表达一种IgG 输送蛋白,称为新生Fc 段受体(neonatal FcR,FcRn) 。 lgG 可选择性与FcRn 结合,从而转移到滋养层细胞内, 并主动进入胎儿血液循环中。 lgG 穿过胎盘的作用是一种重要的自然被动免疫机制,对千新生儿抗 感染具有重要意义。另外,分泌型lgA 可被转运到呼吸道和消化道黏膜表面(图4-8 ) , 在黏膜局部免 疫中发挥重要作用。