“免疫”是机体通过区别“自己”和“非己",对非已物质进行识别、应答和予以清除的生物学效应 的总和。 这些非已物质就是抗原。 抗原(antigen , Ag)是指所有能激活和诱导免疫应答的物质,通常指 能被T、 B 淋巴细胞表面特异性抗原受体(TCR 或BCR)识别及结合,激活T、 B 细胞增殖、分化、产生免 疫应答效应产物(特异性淋巴细胞或抗体),并与效应产物结合,进而发挥适应性免疫应答效应的物 质。 理论上抗原可为自然界所有的外源和自身物质,但机体免疫细胞通常识别的抗原是蛋白质,也包 括多糖、脂类和核酸等。
第一节 抗原的性质与分子结构基础
并非所有的外源或自身物质都是抗原,具备免疫原性和免疫反应性两个重要特性的物质才是抗 原。 抗原诱导机体产生的适应性免疫应答仅对该抗原专一,而与其他抗原无关,这一性质称为免疫应 答的抗原特异性。 适应性免疫应答之所以具有抗原特异性,是由千免疫应答是由TCR/BCR 识别抗原 所包含的最小的基本结构单位-抗原表位所诱导的。
一、抗原的基本特性:免疫原性与免疫反应性
抗原具备两个重要特性:免疫原性(immunogenicity) 和免疫反应性(immunoreactivity) 。 免疫原 性指抗原被T、 B 细胞表面特异性抗原受体(TCR 或BCR)识别及结合,诱导机体产生适应性免疫应 答的能力;免疫反应性是指抗原与其所诱导产生的免疫应答效应物质(活化的T/B 细胞或抗体)特 异性结合的能力。 同时具有免疫原性和免疫反应性的物质称为完全抗原(complete antigen) 。 然 而,某些小分子物质单独不能诱导免疫应答,即不具备免疫原性,但当其与大分子蛋白质或非抗原 性的多聚赖氨酸等载体交联或结合后可获得免疫原性,能诱导免疫应答,此类小分子物质称为半 抗原(hapten) , 又称不完全抗原( incomplete antigen) 。 半抗原可与免疫应答效应物质结合, 具备免 疫反应性。
结构复杂的蛋白质大分子通常为完全抗原,许多小分子化合物及药物属半抗原,如青霉素降解产 物青霉烯酸,本身并无免疫原性, 一旦与血清蛋白结合可成为完全抗原,诱导机体产生IgE 抗体并介 导 I 型超敏反应(青霉素过敏)。
二、适应性免疫应答的抗原特异性
抗原诱导的免疫应答具有抗原特异性(antigenic specificity) , 即抗原刺激机体产生适应性免疫应 答及其与应答效应产物发生结合均显示专一性,某一特定抗原只能刺激机体产生针对该抗原的活化 T/B 细胞或抗体,且仅能与该淋巴细胞或抗体发生特异性结合。 特定抗原与特异性T 细胞或特异性 抗体专一结合的特性,是目前免疫学检测、 诊断及治疗技术的分子基础。 如乙型肝炎病毒表面抗原 (HBsAg), 能诱导机体产生HBsAg 特异性抗体,该抗体仅与HBsAg 特异性结合,不会与乙型肝炎病毒 的其他抗原(如核心抗原)或其他病毒抗原发生结合。 利用这一特性研制的人血清HBsAg 检测试剂 盒,可判断是否感染了乙型肝炎病毒。
三、决定抗原特异性的分子结构基础:抗原表位
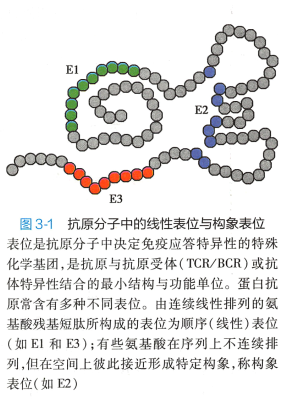
1. 抗原表位的概念 T、 B 细胞通过其表面的特异性抗原受体(TCR/BCR) 对抗原的识别呈现 高度特异性;被抗原活化的T 细胞和活化B 细胞效应产物抗体与抗原的结合也呈高度特异性。 上 述两种特异性的分子基础取决于抗原分子所含的抗原表位(epitope) , 又称抗原决定基(antigenic determinant) 。 表位是抗原分子中决定免疫应答特异性的特殊化学基团,是抗原与T/B 细胞抗原受 体(TCR/BCR) 或抗体特异性结合的最小结构与功能单位。 表位通常由5 -15 个氨基酸残基组成, 也可由多糖残基或核昔酸组成。 1 个抗原分子中能与抗体结合的抗原表位总数称为抗原结合价 (antigenic valence) 。 天然蛋白大分子通常为多价抗
原,含多种多个抗原表位,可诱导机体产生含有多种 特异性抗体的多克隆抗体。 一个半抗原相当于一个抗 原表位,仅能与TCR/BCR 或抗体分子的一个结合部位 结合。
2. 抗原表位的类别 根据抗原表位中氨基酸的 空间结构特点,可将其分为顺序表位(sequential epitope) 和构象表位(conformational epitope) (图 3 -1) 。 顺序表位由连续线性排列的氨基酸构成,又称 线性表位(linear epitope) ; 而构象表位由不连续排 列、但在空间上彼此接近形成特定构象的若干氨基 酸组成。
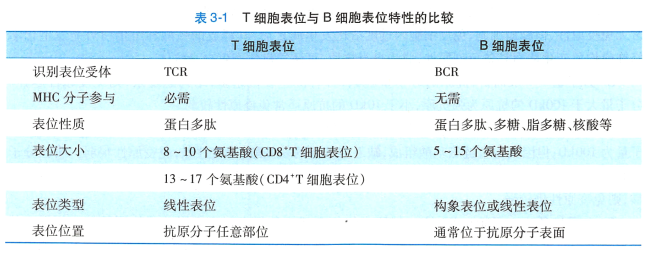
根据T、B 细胞所识别的抗原表位的不同,表位可 分为T 细胞表位和B 细胞表位。 T 细胞仅识别由 APC加工后与MHC 分子结合为复合物并提呈于APC 表面 的线性表位,此类表位称T 细胞表位。 T 细胞表位又 可分两种: (1)CD8+T 细胞识别的表位,含8 -10 个氨基酸,其中第2 、9 位氨基酸为铀定氨基酸(anchor residue) ; (2)CD4 +T 细胞识别的表位,较长,含1 3 -17 个 氨基酸。 BCR 或抗体识别的B 细胞表位,无需APC 加工和提呈,含5 -15 个氨基酸,多为构象表位, 少数为线性表位,位于抗原分子表面。 表3-1 是T 细 胞表位和B 细胞表位特性的比较。
四、半抗原载体效应
天然蛋白抗原同时存在T 和B 细胞表位,可分别激活T 细胞和B 细胞,其中B 细胞激活有赖于T细胞辅助。 某些人工合成的简单有机化学分子属半抗原,免疫原性很低,须与蛋白质载体偶联才可诱 导抗半抗原的抗体产生。 其机制为:B 细胞特异性识别半抗原; 蛋白载体含CD4+T 细胞表位,被B 细 胞或其他APC 提呈并活化CD4+T 细胞。 由此,T-B 细胞通过载体而相联系,Th 细胞借此相互作用辅 助激活B 细胞。
五、共同抗原表位与交叉反应
某些抗原分子中含多个抗原表位,而不同抗原间可能含相同或相似的抗原表位,称为共同抗原表 位(common epitope) 。 因此,某些抗原诱生的特异性抗体或活化淋巴细胞,不仅可与自身抗原表位特 异性结合,还可与其他抗原中相同或相似的表位反应,此为交叉反应(cross-reacti on) 。 含共同抗原表 位的不同抗原称为交叉抗原(cross antigen)。 机体感染链球菌导致 风湿性心脏病的主要原因是链球菌中含有与心肌抗原的交叉抗原,其诱导的抗体与T 细胞可交叉攻 击心肌。

