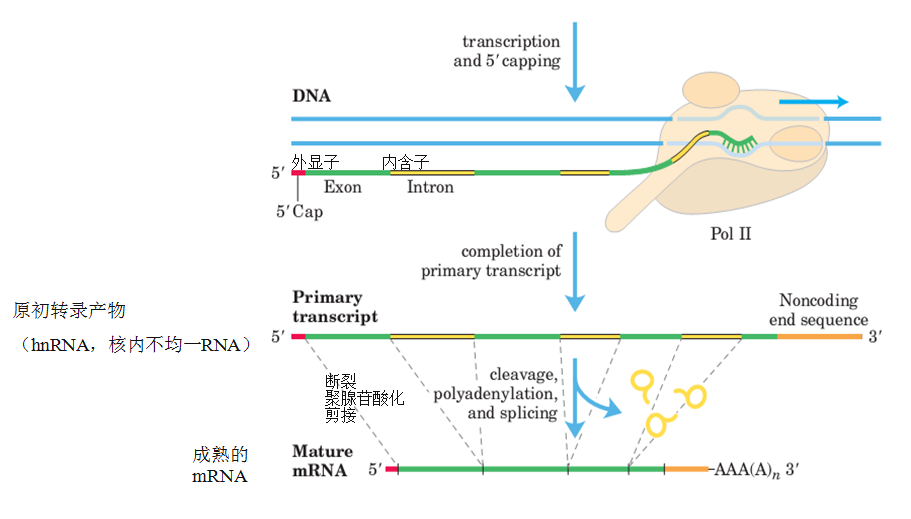
一、mRNA的转录后加工
(1)5’端加帽子结构
(2)3’端加poly(A)尾巴结构
(3)mRNA的剪接
1.首尾的修饰
(1) 5´-端加帽:m7GpppG——
(2) 3´-端加尾:多聚腺苷酸 (poly A)

2 .帽子结构 5’Capping
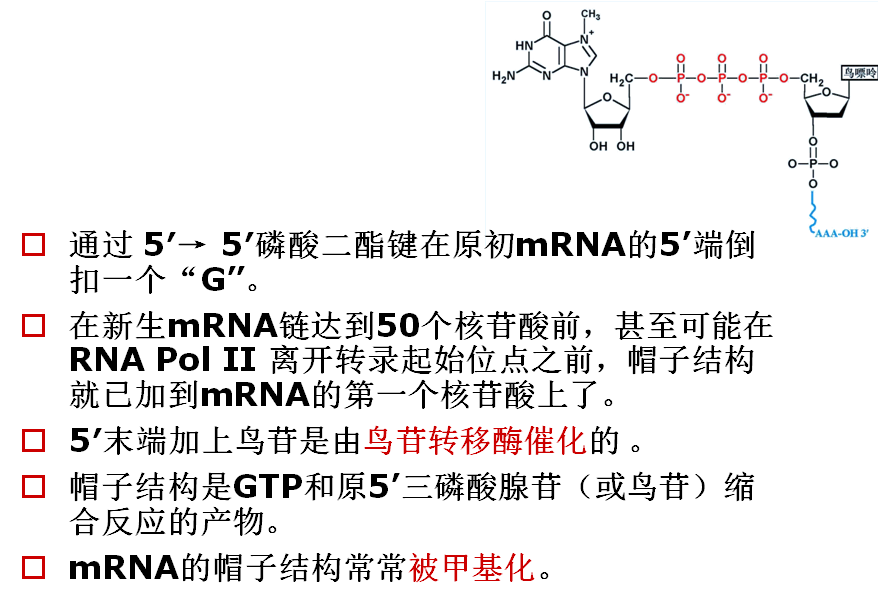
3.帽子结构的生成
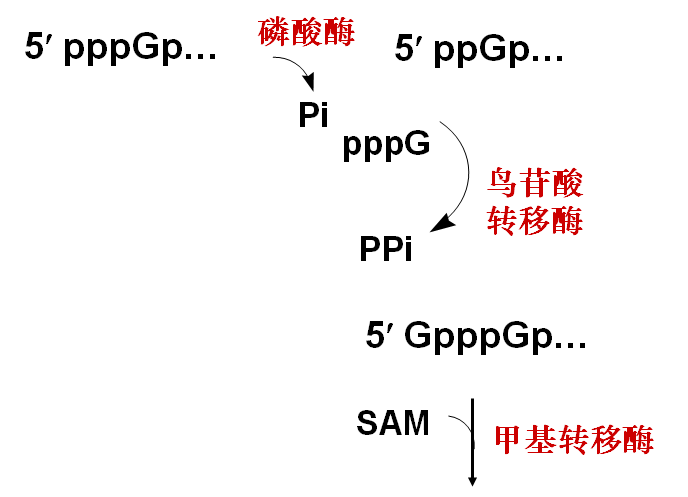
加帽过程

二、真核生物mRNA的“帽子”结构
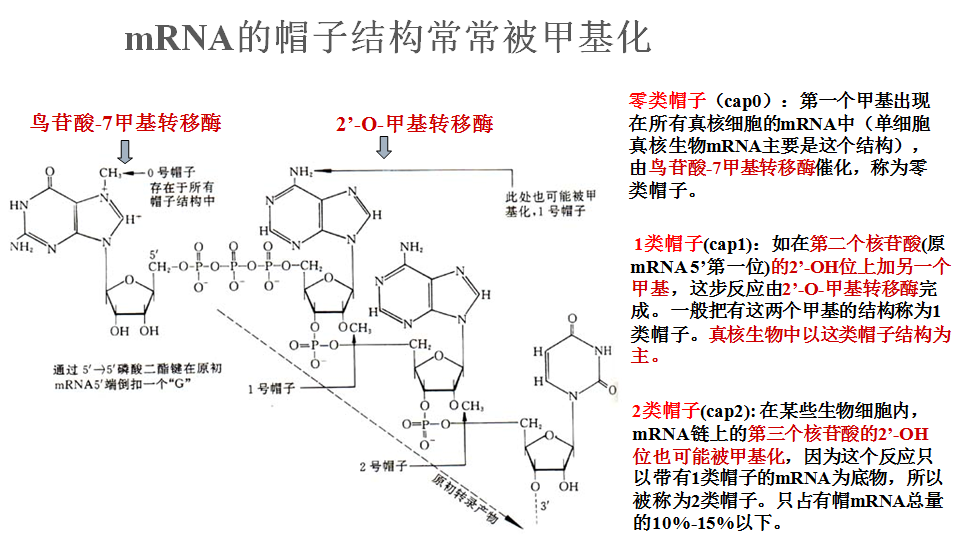
加帽的功能
mRNA 5’加帽的功能主要表现在4个方面:
(1)阻止mRNA的降解:细胞内存在许多RNA酶,它们可从5’端可攻击游离的RNA分子。当mRNA的5’端加上m7GpppG帽子后,可阻止RNase的切割,延长mRNA的半衰期。
(2)提高翻译效率 真核生物mRNA必需通过5’帽结合蛋白才能接触核糖体,起始翻译。缺少加帽的mRNA由于不能被5’帽结合蛋白识别,其翻译效率比加帽的mRNA低20倍。
(3)作为进出细胞核的识别标记 凡由RNA聚合酶II转录的RNA均在5’端加帽,包括snRNA,这是RNA分子进出细胞核的识别标记。如:U6 snRNA 由RNA聚合酶III转录,其5’端保留3个磷酸基团,无帽子结构,因而不能输出细胞核。
(4)提高mRNA的剪接效率 5’帽结合蛋白涉及第一个内含子剪接复合物的形成,直接影响mRNA的剪接效率。
1. 3´-端加尾:多聚腺苷酸 (poly A)
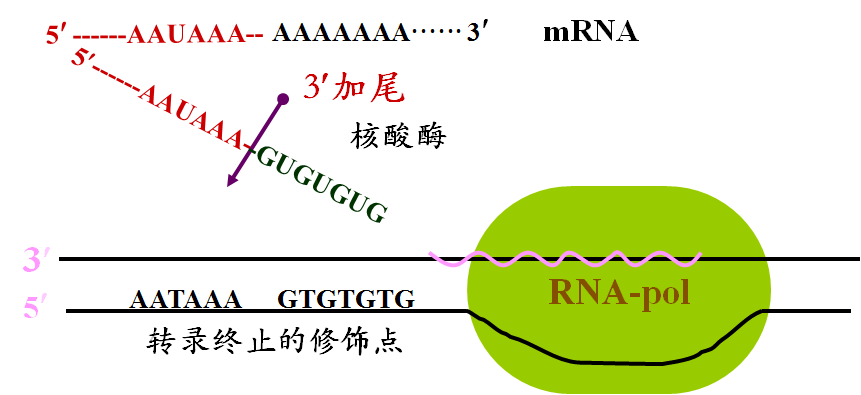
绝大多数真核生物mRNA具有多聚(A)尾巴。
除组蛋白基因外,真核生物mRNA的3’末端都有多聚(A)序列,其长度因mRNA种类不同而变化,一般为40-200个左右。
由多聚(A)聚合酶催化的;
它是在转录后加上的;
Poly(A)被特异的蛋白质PABP结合。
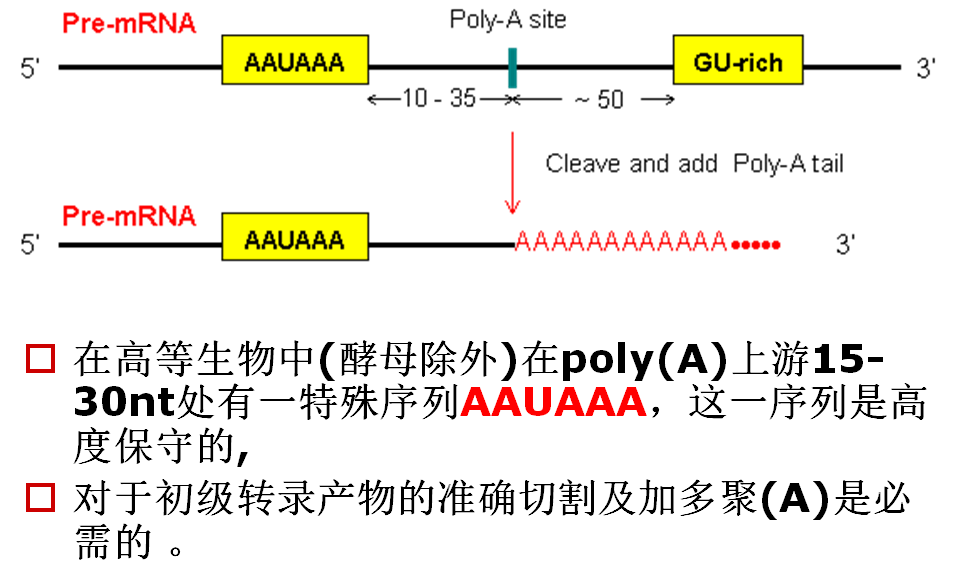
2.真核生物mRNA中的加多聚A反应
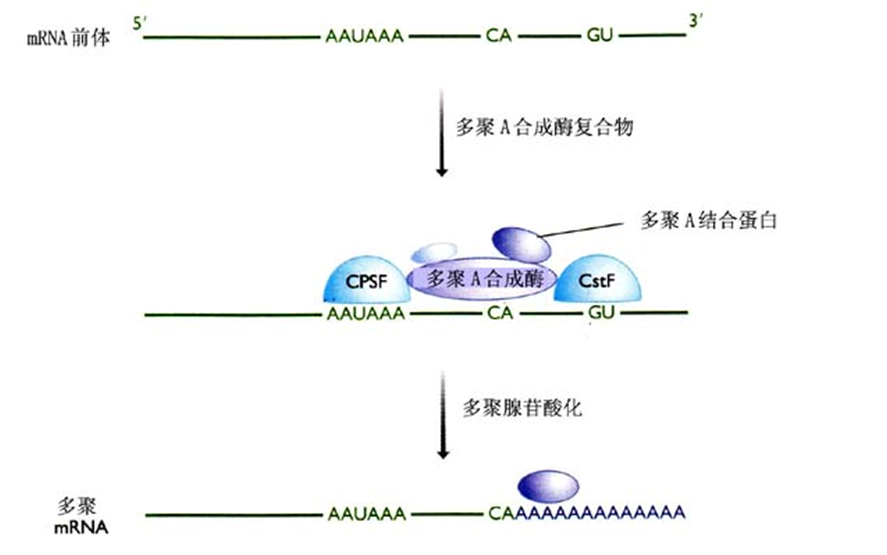

三多聚腺苷化(polyadenylation) 反应要经过2个阶段
(1)首先将一个短的寡聚A序列(10nt)加到3’端,
此反应绝对依赖于AAUAAA序列,这是由poly(A)聚合酶在特殊因子指导下完成的。
(2)寡聚A尾巴延伸到240nt的长度。
此反应并不需要AAUAAA序列,但需要一个识别寡聚A并指导poly(A)聚合酶延伸的刺激因子。
四、多聚(A)的功能
1.是mRNA由细胞核进入细胞质所必需的形式;
2.mRNA刚从细胞核进入细胞质时,其多聚(A)尾巴一般比较长,随着mRNA在细胞质内逗留时间延长,多聚(A)逐渐变短消失,mRNA进入降解过程。
3.它大大提高了mRNA在细胞质中的稳定性。
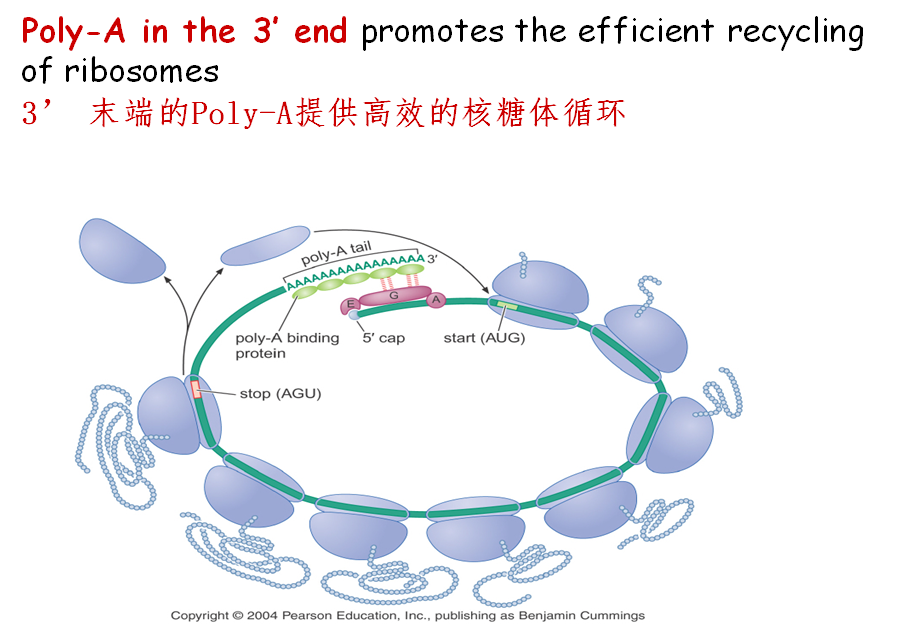
4.它可促进核糖体的有效循环。
1.mRNA的剪接
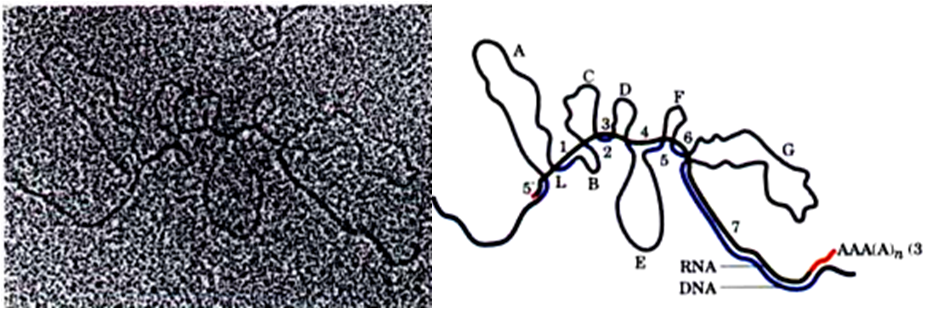
(1). hnRNA
核内的初级mRNA称为核内不均一RNA (hnRNA)
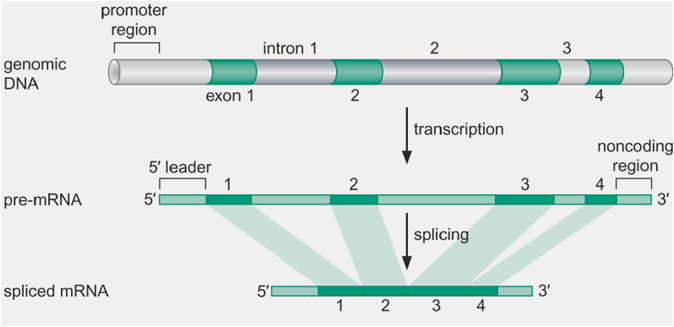
2.断裂基因(splite gene)、外显子(exon)和内含子(intron)
断裂基因(splite gene):真核生物结构基因内部,若干编码区和非编码区相间隔镶嵌存在,称为断裂基因。
外显子(exon):在断裂基因中原初转录产物上出现,并表达为成熟RNA的核酸序列。
内含子(intron):隔断基因的线性表达,而在原初转录产物剪接过程中被除去的核酸序列。

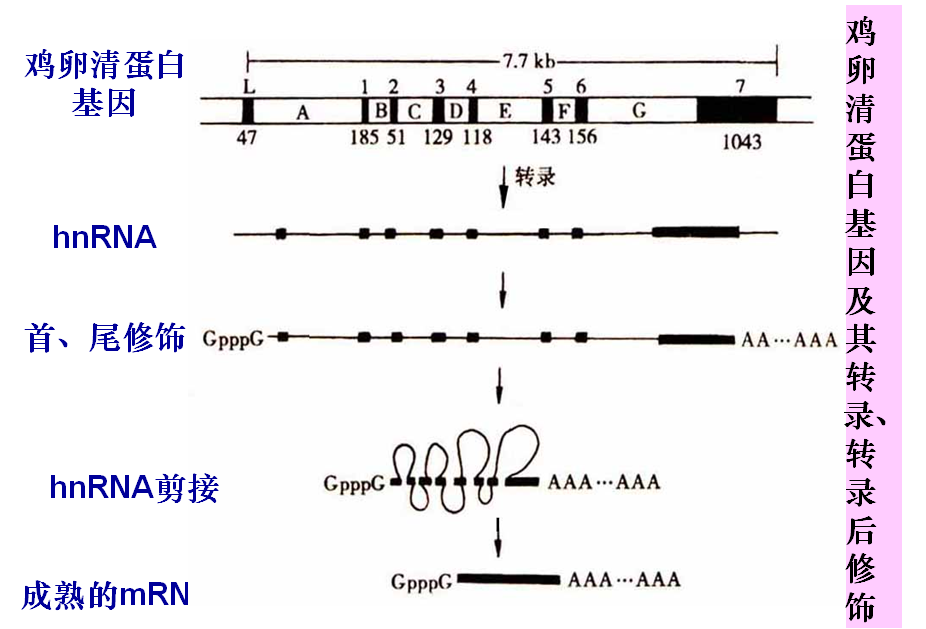
3. mRNA的剪接
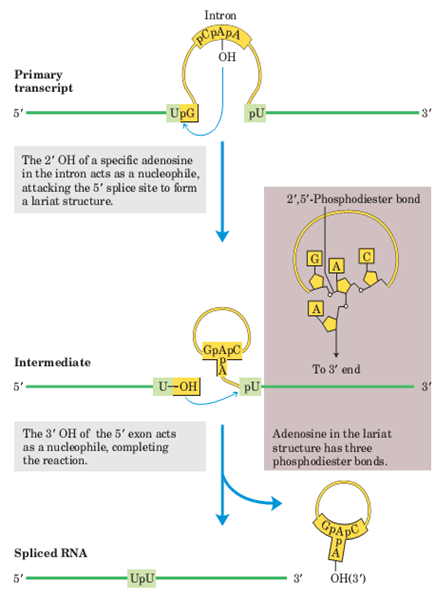
—— 除去hnRNA中的内含子,将外显子连接。
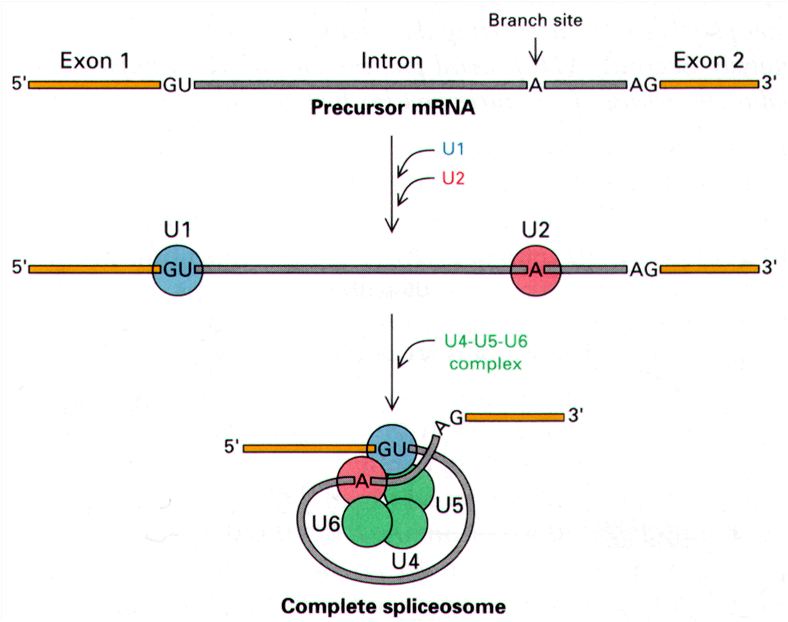
snRNA (small nuclear RNA)
剪接体:是由核内U系5种snRNA(U1、U2、U4、U5、U6)和20多种剪接的蛋白因子构成,催化核内mRNA前体的剪接。

4.I类内含子的自我剪接过程

I类内含子的自我剪接过程(动画)

5.II类内含子的自我剪接过程
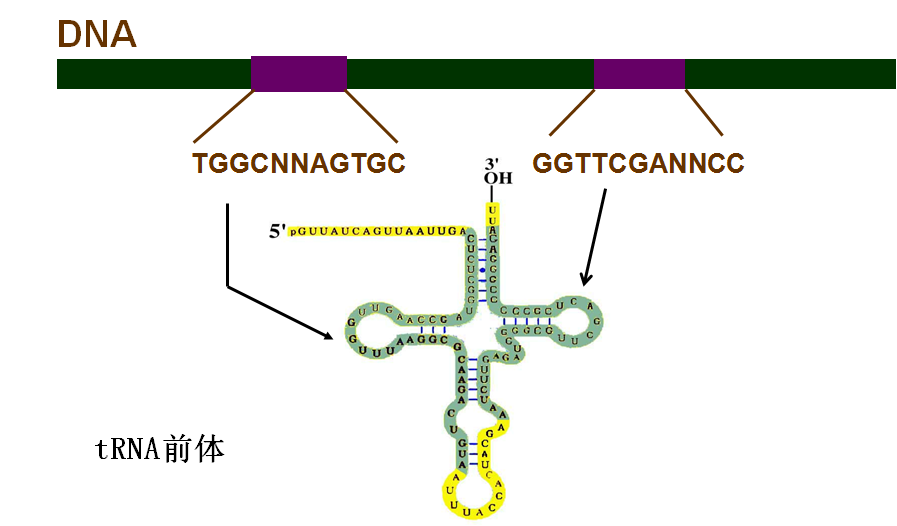
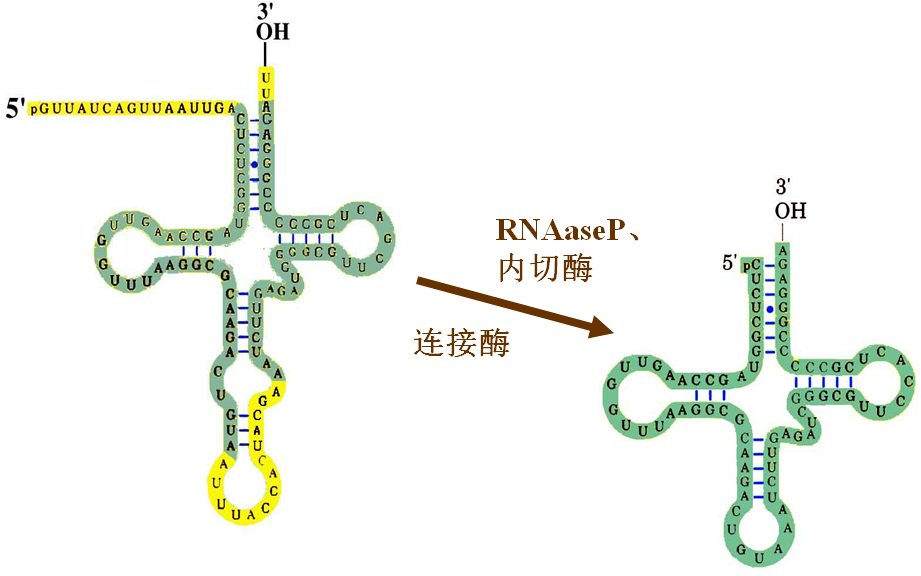
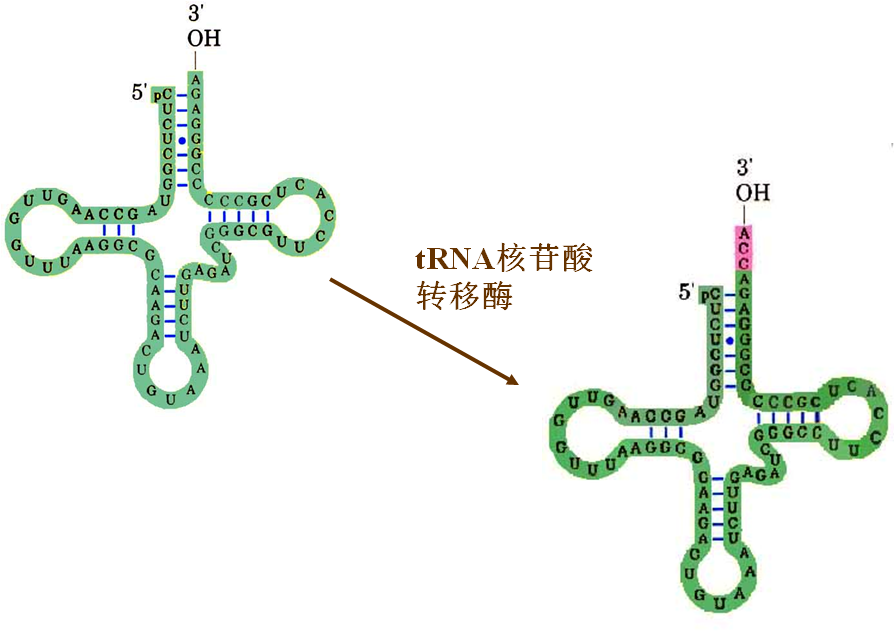
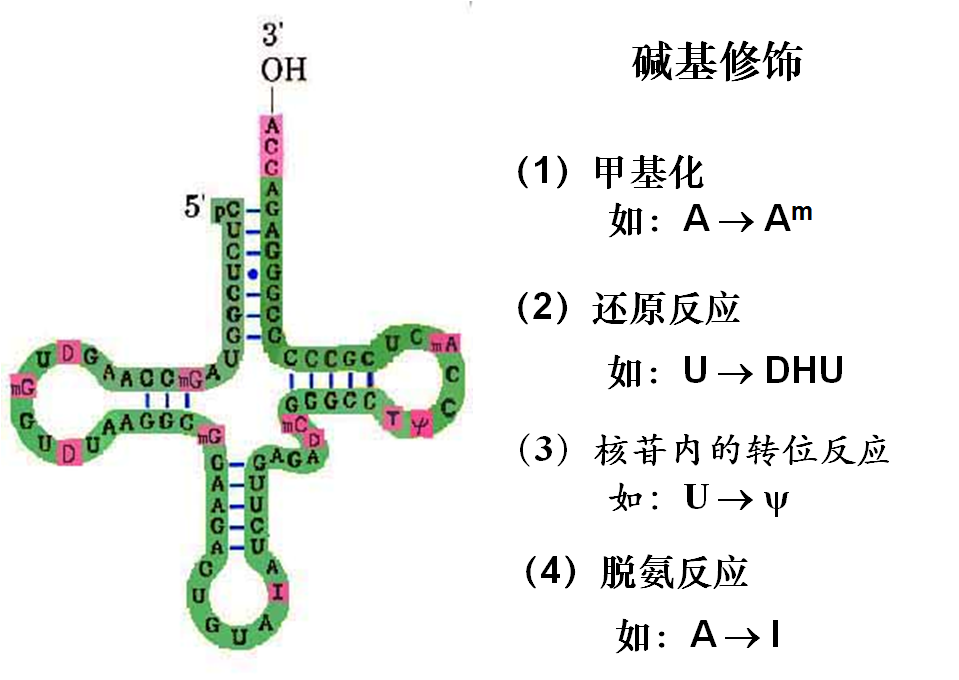
五、tRNA的转录后加工
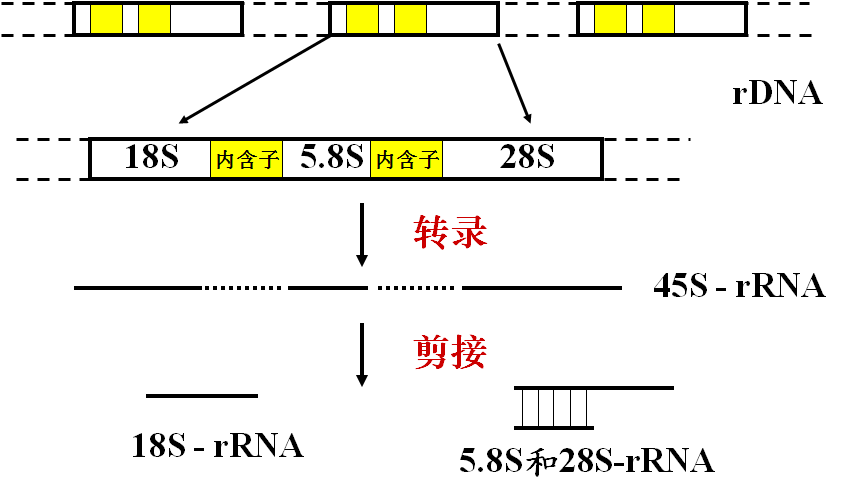
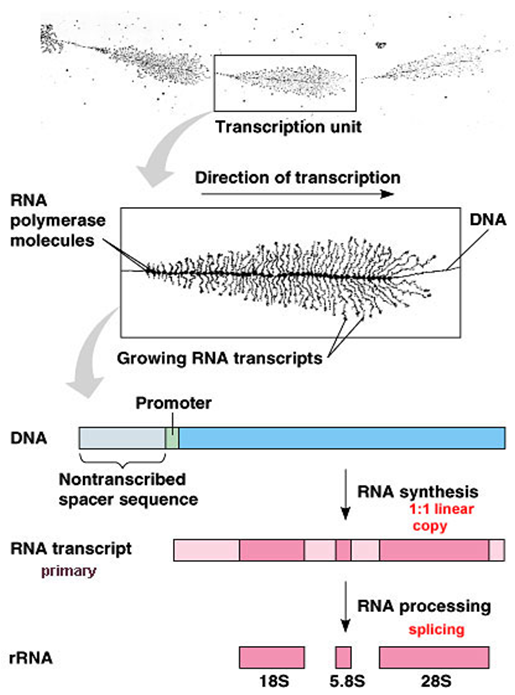
六、rRNA的转录后加工