
一、转录单元(transcription unit)
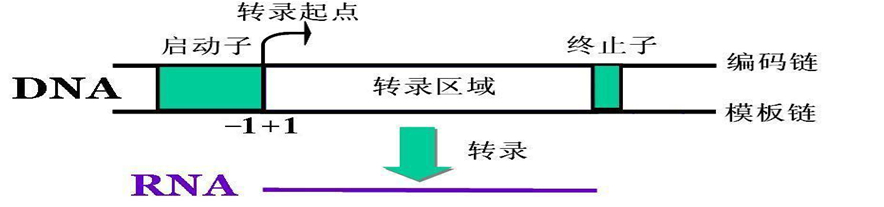
转录单元(transcription unit):RNA链的转录起始于DNA模板的一个特定起点(启动子),并在一终(转录单位) 点处(终止子)终止,此转录区域称为转录单位。一个转录单位可是一 个基因,也可是多个基因。
启动子(promoter) :指RNA聚合酶识别﹑结合和开始转录的一段DNA序列。其确保转录精确有效地起始。
启动子是一段位于结构基因5’端上游区的保守的DNA序列,能活化RNA聚合酶,使之与模板DNA准确地相结合并具有转录起始的特异性。
二、启动子与转录起始
大肠杆菌RNA聚合酶与启动子的相互作用主要包括:
1.启动子区的识别;
2.酶与启动子的结合;
3.σ因子的结合与解离。
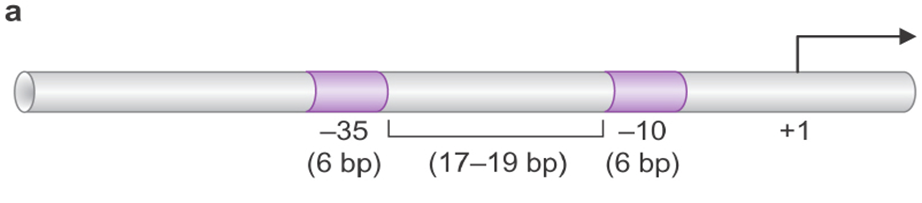
三、原核生物的启动子
-10区(Pribnow区)的发现
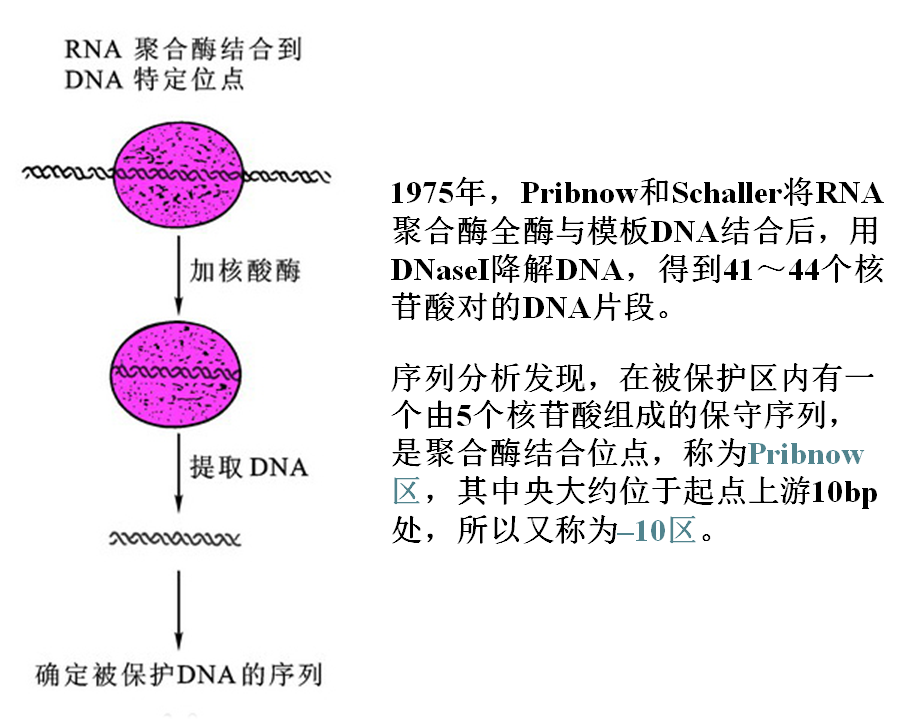
2.-10序列 (Pribnow框盒)

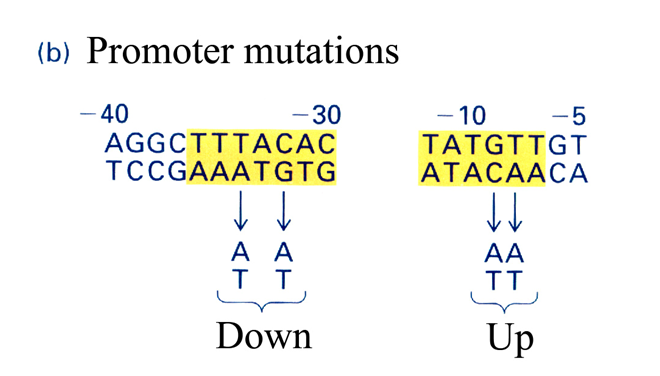
如果把Pribnow区从TATAAT变成AATAAT就会使该启动子发生下降突变(down mutation);
如果增加Pribnow区的共同序列,将乳糖操纵子的启动子中的TATGTT变成TATATT,就会提高启动子的效率,称为上升突变(up mutation)。
3.-35区的发现

科学家又从噬菌体的左、右启动子PL及PR和SV40启动子的–35 bp附近找到了另一段共同序列:TTGACA。
4.-35序列 (Sextama盒)
其保守序列为TTGACA,
与-10序列相隔16-19bp。
功能:
(1)为RNA pol的识别位点。
(2)RNA Pol的核心酶只能起到和模板结合和催化的功能,并不能识别-35序列,只有σ亚基才能识别-35序列,为转录选择模板链。
5.100个E.coli的不同启动子的序列测定结果
–35区 –10区
……T85T83G81A61C69A52……T89A89T50A65A100……
大部分启动子都存在这两段共同序列,即位于–10 bp处的TATA区和–35 bp处的TTGACA区。
它们是RNA聚合酶与启动子的结合位点,能与σ因子相互识别而具有很高的亲和力。
6.-10区和-35区的间隔序列

7.Some stronger bacterial promoter

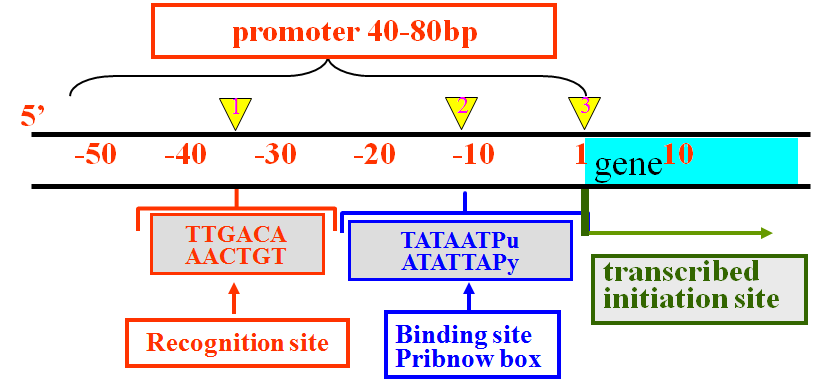
(1)结构典型,都含有识别(R),结合(B)和起始(I)三个位点;
(2)序列保守,如-35序列,-10序列结构都十分保守;
(3)位置和距离都比较恒定;
(4)直接和多聚酶相结合;
(5)常和操纵子相邻;
(6)都在其控制基因的5′端;
(7)决定转录的启动和方向。
原核生物启动子 的共同的特点
四、真核生物的启动子
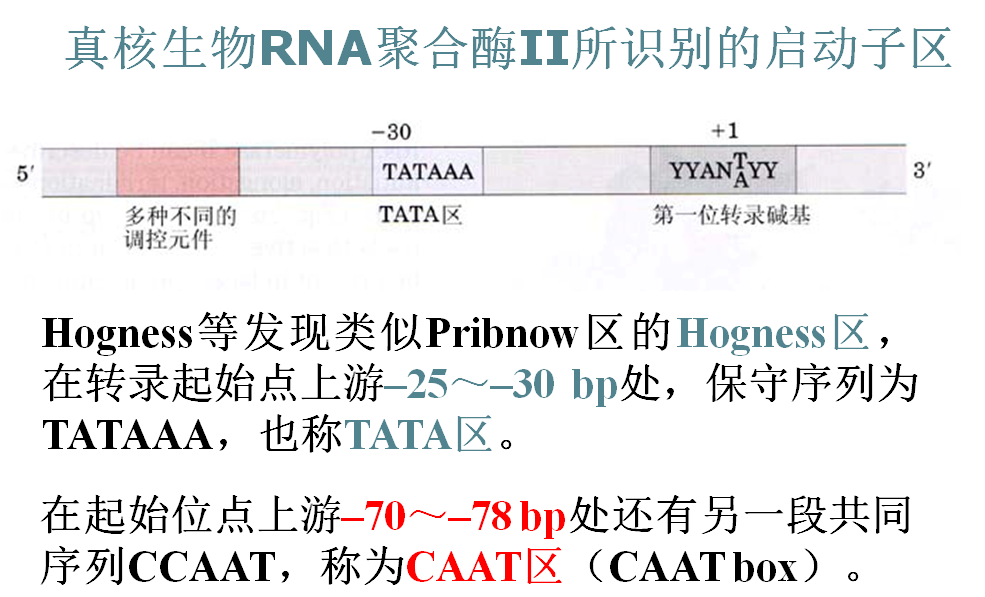
真核基因的启动子


2.真核生物启动子对转录的影响
(1)TATA区--使转录精确地起始:
如果除去TATA区或进行碱基突变,转录产物下降的相对值不如CAAT区或GC区突变后明显,但发现所获得的RNA产物起始点不固定。
(2)CAAT区和GC区主要控制转录起始频率:基本不参与起始位点的确定。
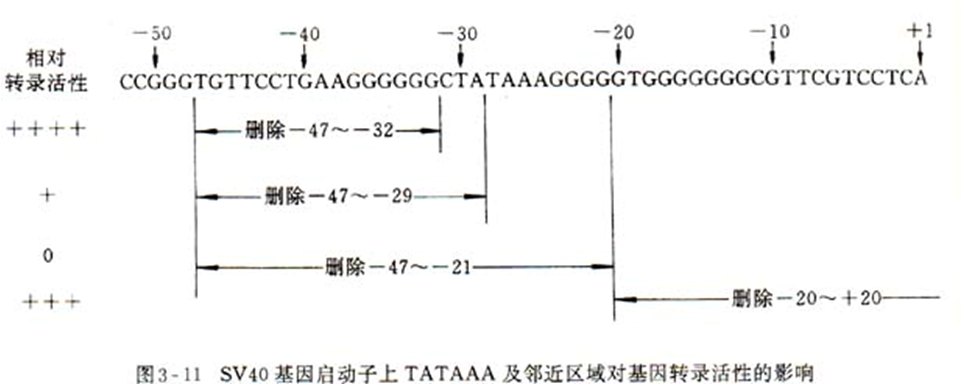
研究SV40晚期基因启动子发现,上游激活区的存在与否,对该启动子的生物活性有着根本性的影响。若将该基因5' 上游–21~–47核苷酸序列切除,基因完全不表达。
远端调控区
增强子(enhancer)
1981年由研究人员发现,又称远上游序列(far upstream sequence)。
增强子(enhancer 或称强化子):一种能强化转录起始的序列,其不是启动子的一部分,但能促进转录起始,除去之则基因转录水平大大降低。
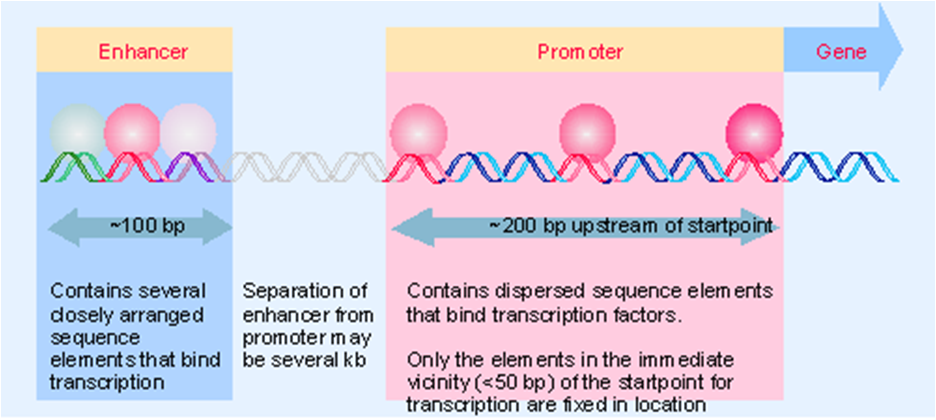
增强子很可能通过影响染色质DNA-蛋白质结构或改变超螺旋的密度而改变模板的整体结构,从而使得RNA聚合酶更容易与模板DNA相结合,起始基因转录。
3.增强子特点
① 具有远距离效应。
常在上游-200bp处,但可增强远处启动子的转录,即使相距十几Kb也能发挥其作用;
② 无方向性。
无论在靶基因的上游,下游或内部都可发挥增强转录的作用;
③ 顺式调节。
只调节位于同一染色体上的靶基因,而对其它染色体上的基因无作用;
④ 无物种和基因的特异性,
可以接到异源基因上发挥作用;
⑤ 具有组织的特异性。
SV40的增强子在3T3细胞中比多瘤病毒的增强子要弱,但在HeLa细胞中SV40的增强子比多瘤病毒的要强5倍。增强子的效应需特定的蛋白质因子参与。
⑥ 有相位性。其作用和DNA的构象有关。
⑦ 有的增强子可以对外部信号产生反应。
如热体克基因在高温下才表达。编码重金属蛋白的金属硫蛋白基因在镉和锌存在下才表达。某些增强子可以被固醇类激素所激活。
4.真核生物的启动子特点

(1)有多种元件:TATA框,GC框,CATT框,OCT等;
(2)结构不恒定。有的有多种框盒,如组蛋白H2B;有的只有TATA框和GC框,如SV40早期转录蛋白;
(3)它们的位置、序列、距离和方向都不完全相同;
(4)有的有远距离的调控元件存在,如增强子;
(5)这些元件常常起到控制转录效率和选择起始位点的作用;
(6)不直接和RNA pol结合。转录时先和其它转录激活因子相结合,再和聚合酶结合。

