
电生理学发展简史
生物电活动是机体一种基本的生命现象,它产生的基础是细胞膜上离子通道活动的总和效应。从生物电现象的发现到如今对离子通道功能与结构如此深入的了解,电生理学走过了 200 多年的历程。
一、生物电现象的发现
最初的实验研究是从18世纪后叶开始的。当时没有任何测量电流的仪器,只是发现利用电容器(如雷顿瓶)的放电,或雷电发生时竖起一根长导线,引导大气中的电,都可以刺激蛙的神经肌肉标本,引起肌肉收缩,所以当时就用蛙的神经肌肉标本作为电流存在的标志。 1791年意大利解剖学 教授 Galvani L 发现,如果将蛙腿的肌肉置于铁板上,再用铜钩钩住蛙的脊髓,当铜钩与铁板接触时肌肉就会发生收缩。他把这个现象的发生归因于机体的 “ 动物电 ” ( animal electricity )。他认为神经与肌肉带有相反的电荷,肌肉带正电,神经带负电,金属导体的作用是把神经与肌肉之间的电路接通。同时代的意大利物理学家 Volta A 不同意 Galvani 的见解,他认为实验中发现的电现象,不是动物机体产生的动物电,而是由于实验中连接肌肉和神经的金属不同所致,是不同金属接触时产生的电流刺激了肌肉标本,如果用同一种金属作导体,收缩就不会发生。事实上, Volta 和 Galvani 的观点都有其正确的一面。 Volta 后来因此而发明了伏特电池; Galvani 则继续进行了一个出色的实验。在无金属参与的情况下,他将一个肌肉标本横断,又将另一个神经肌肉标本的神经干搭在横断肌肉上,并使之跨越肌肉的完好面和损伤面,结果该神经支配的肌肉产生收缩,证实了动物电的存在。这成为第一次观察到生物电存在的电生理实验。但是直接测量到生物电的实验是在电流计发明之后。
1825 年意大利物理学家 Nobili 发明电流计。
1837年意大利物理学教授 Matteucci C 用电流计在肌肉的横断面与未损伤部位之间,测量到电流流动,电流是从未损伤部位流向横断面的,所以横断面呈负电位。这是第一次直接测量到生物体内存在生物电的实验。
1843 年瑞士生理学家 Du Bios-Reymond 用电流计观察到神经的损伤电位,也是损伤部位呈负性。 1849 年,他又发现神经在活动期间出现负波动,即使用电流计从细胞外记录到的动作电位。
1850 年 von Helmholtz H 测定了神经传导速度,证明蛙神经的传导速度仅 20 ~ 30m/s 。此前人们认为既然电的传导速度等于光速,因而神经的传导速度可能也是光速。
二、早期对生物电发生机制的认识
1. Bernstein 的膜学说
对于这种生物电现象的解释,当时提出了不同的学说。 Du Bios-Reymond 认为,组织内带负电,外表带正电,是正常状态下存在的,即所谓 “ 现存学说 ” ( preexistence theory );而他的学生 Hermann ( Du Bios-Reymond )则认为,组织内的负电是被切割时组织损伤变质造成的,即所谓 “ 变质学说 ” ( alteration theory )。 1890 年,著名的化学家 Ostwald W 提出了膜的通透性理论,即如果在电解质弥散的途径上有一层半透膜,它只允许一种离子通过,而带有相反电荷的另一种离子不能通过,就会通过静电作用限制透过膜的离子不能进一步弥散,如 此 ,在膜两侧就会形成电位差,它的大小可按 Nernst 公式计算。 1902 年 Du Bios-Reymond 的另一名学生 Bernstein J 接受了 Ostwald 通透性理论,在现存学说的基础上提出了“ 膜学说 ”( membrane theory )。他根据细胞内液比细胞外液含较多的K+ ,而细胞损伤处电位较完好处为低的事实,推测静息时细胞内电位低于细胞外,并假定静息时细胞膜只K+有通透性,由于胞内带正电荷的K+顺浓度差扩散到膜外,相应的负电荷仍留在膜内,使细胞膜呈现外正内负的极化状态,形成静息电位。按照 Bernstein 的设想,细胞的静息电位就等于 K+的平衡电位。动作电位则是由于膜在一瞬间失去了对 K+的选择性通透,变得对所有离子通透性一过性地升高,导致膜两侧电位差瞬间消失。 1904 年,他又设计了一个精巧的实验,证实肌肉切断后断面的负电位在 0.3 s 后即出现,并持续缓慢地减小而不是逐渐增大,从而用事实否定了变质学说。因此膜学说在此后 30 多年的时间内为多数人所接受,不过它却一直未能得到实验证实。这是因为要想测量膜两侧的电位差,必须要将一个电极插入细胞内,这就要求插入的记录电极直径很细,不能损伤细胞;插入处也不能漏电。显然,这种技术上的限制在当时是很难克服的。
2. 细胞跨膜电位的记录
1936 年,生物学家 Young JZ 发现了头足类软体动物枪乌鲗的巨大神经轴突,其直径可达 1mm 。这与最大直径不超过 20μm 的脊椎动物神经纤维相比,无疑是研究跨膜电位的绝好材料。 1939 年,英国生理学家 Hodgkin AL 和 Huxley AF 将直径为 0.1mm ,内部充满海水的毛细玻璃管纵向插入枪乌鲗大神经轴突(直径 0.5mm )的断端,作为细胞内电极,而将另一电极置于浸泡细胞的海水中,于是在毛细管尖端和细胞外电极之间记录到约 60mV 的电位差,胞 质 为负。将轴浆内电极靠近膜内侧或向中心移动,电位差均不变,表明电位差存在于膜的两侧。这样,他们便利用枪乌鲗巨大轴突首次记录到膜两侧的静息电位。这一工作的意义在于实验测定的静息电位与根据细胞膜内、外钾浓度经 Nernst 公式计算的钾平衡电位( - 75mV )非常接近,从而有力地支持了 Bernstein 关于静息电位状态下细胞膜选择性对钾离子有通透性的膜学说。但是,正如在图 1 中所看到的,实测的静息电位值总是略小于 K + 平衡电位,这说明细胞膜并不是像原来设想的那样只对 K + 有通透性,而可能对其他离子,特别是Na + 亦有一定的通透性。
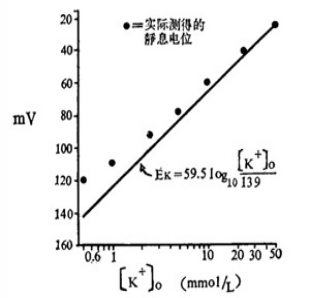
图1 细胞外 K + 浓度对蛙缝匠肌静息电位的影响
图中的圆点是不同 K + 浓度时静息电位的实测值;直线是根据 Nernst 公式的计算值
膜学说关于动作电位产生机制的解释也得到膜阻抗测量实验的支持。 1939 年, Cole KS 和 Curtis HJ 利用惠斯登电桥( Wheatstone bridge )观测枪乌鲗巨 大 轴突动作电位期间膜电阻和膜电容的变化,发现动作电位期间膜电容基本不变,而膜阻抗显著降低(膜电导增大),由静息时的 1000Ωcm 2 下降到 25Ωcm 2 ,支持动作电位期间膜对多种离子通透性的增大。
但是,Hodgkin 和 Huxley 利用细胞内电极在记录枪乌鲗巨轴突静息电位的基础上观测受刺激后产生的动作电位时,却使人意外地发现,动作电位的幅度大大超过了零电位( + 30 ~+ 50mV ),出现膜电位的倒转(超射),这是用膜对各种离子通透性普遍增大的理论无法解释的。考虑到经公式计算的钠平衡电位在 +50mV至+70mV , Hodgkin 和 Huxley 提出了一个新的 “ 离子学说 ” ,即动作电位期间膜对钠通透性瞬间增大并远远超过了对钾的通透性。
3. Hodgkin 的离子学说
1949 年 Hodgkin 和 Huxley 用氯化胆碱或葡萄糖等张地替代神经纤维周围海水中的 NaCl ,结果表明,这种状况下的动作电位幅度、升支上升速度和动作电位传导速度都下降了,其下降程度随钠离子被替代比例的增大而增大(图 3 ),从而证实了他们的离子学说,即安静时选择性地对钾通透,而兴奋时对钠的通透性瞬间增大。 1951 年,Keynes 和 Lewis 利用同位素 24 Na+ 的定量研究也证明,在给予枪乌鲗神经巨大轴突几分钟连续刺激后,浸浴液中的 24 Na+大量进入轴浆,经计算,每次动作电位进入细胞内的 Na+ 约为 21000/ mm 2 。如果细胞膜的膜电容为 1 m F/cm 2 ,这一Na+流量足以使膜去极化达 100mV 以上。不过这种实验还不能分析动作电位期间的 Na+ 内流和膜对 Na+ 通透性变化的精确的时间过程。
1949 年 Ling 和 Gerard 创立了微电极技术,使人们有可能对多种细胞作细胞内电位的直接测量。各类可兴奋细胞上所测得的静息电位与动作电位大致与上述情况相似 。
4. 探讨动作电位期间膜对离子通透性的变化 ―― 电压钳技术的建立及其贡献
细胞膜对动作电位期间离子通透性(膜电导)变化的直接测定是从应用电压钳( voltage clamp )技术以后开始的。该技术是 1949 年由 Cole KS 及 Marmont G 设计的,后经 Hodgkin 和 Huxley 和 Katz 等加以改进,并成功地应用于枪乌 鲗 巨轴突动作电位期间离子电流的研究。电压钳技术的基本原理就是欧姆定律,即 :
I m = g·V m
式中 g 为膜电导, V m 为膜电位, I m 为膜电流。该式表明,为了测定动作电位期间膜电导 g 的变化,只要膜电位 V m 保持不变,膜电流 I m 就可以作为观测膜电导 g 变化的指标。但事实上,动作电位期间 V m 发生了快速变化,使膜电导 g 与膜电位 V m 互为因变量,又夹杂产生了大量的电容电流,从而使 I m 的测量失去意义。电压钳技术就是为了克服这一困难而设计的。它通过一个反馈电路向膜内注入电流迫使 V m 保持恒定,此时 I m 的变化就可反映 g 的变化。
Hodgkin 和 Huxley 利用电压钳技术,直接测定了动作电位期间的膜电流,推算出钠电导和钾电导的变化,并用一组数学模型方程式,定量地描述了这个变化过程,很好地模拟了包括动作电位形状在内的可兴奋膜的各种兴奋与传导的特性。他们提出的描述电压门控通道门控动力学的基本概念至今仍被沿用。他们也因此荣获 1963 年度诺贝尔生理学 或医学奖。
三、离子通道概念的提出
1955 年, Hodgkin 和Keynes 首次提出细胞膜上含有离子通道(ion channel ),它们分别对某种离子有选择性的通透能力,并且通过自己的“开放” 或“关闭”等状态的改变而影响和决定膜对某种离子的通透性。此后,陆续发现一些毒物或药物能够选择性地阻断膜对某些离子的通透,如河豚毒可以选择性地阻断膜对Na+的通透而不影响对K+的通透,四乙铵则可影响膜对K+ 的通透而不影响对Na+的通透。这些毒物或药物的作用很可能就是特异地结合了膜上与某种离子通透有关的通道结构。有人用同位素标记的河豚毒做实验,发现它们只和细胞膜上散在的一些蛋白质分子作 1:1 的结合,并可由此算出 Na+通道的密度。 Na+通道在枪乌鲗巨大轴突膜上的密度约为每平方微米550个,在兔迷走神经纤维膜上约为 100 个,在一些有髓鞘神经纤维朗飞结处的膜上可达104~105个。如果把计算所得的 Na+通道数目和膜兴奋时的 Na + 内流量作比较,则可得到兴奋时每秒钟将有多于 107 个 Na+ 流过 Na+ 通道。这个速率超过钠泵主动转运Na+速度的 105倍,比体内一般酶反应的转换速率快 100 倍。再加上此速率的温度系数较低,Q10(即温度每增加 10 度速率增加的倍数)与Na+在水中自由扩散系数的 Q 10 相近似,因此可以设想,当膜对 Na+的通透性增加时,膜上的钠通道蛋白结构中打开了某种水相孔洞。因此,如何能观测到这种推测中的单个离子通道的活动,以证实膜上离子通道的存在,成为当时许多研究者追求的目标。后来膜片钳技术的建立圆满地解决了上述问题,但是在此之前有两个技术曾为人们认识单通道活动的特征和启发膜片钳技术的思路 做 出了贡献,它们是波动分析 ( fluctuation analysis ) 技术和人工脂膜( artificial lipid bilayers )技术。
四、单通道电流的记录 ―― 膜片钳技术的建立和发展
1969 年 Bean 等将电压钳技术应用于塑料隔板小孔上形成的小片人工脂膜,他们发现在几乎绝缘的人工脂质双层中掺入少量的细菌蛋白后,脂膜成为可导电的,并记录到断续的阶梯状的跨膜电流。 1970年 Hadky和Haydon 在同样的人工脂膜中掺入少量的短杆菌肽( gramicidin ),也记录到形式相同的跨膜电流(图 5 )。这些证据表明,阶梯样电流是由于搀入了能形成孔洞样结构的蛋白质所致,每一阶梯反映了单个通道蛋白分子的开放和关闭。因此,单通道的电活动是首先在人工脂膜上记录到的。
参 考 资 料
1. 贾宏钧 , 王钟林 , 杨期东主编. 离子通道与心脑血管疾病. 北京:人民卫生出版社 ,2001
2. Keynes RD , Aidley DJ. Nerve and Muscle. 3rd e dition, Cambridge University P ress, 2001
3.刘振伟编著 . 实用膜片钳技术. 北京 :军事医学科学出版社 , 2006
(吴博威 )

