
第二节 细胞周期和调控
一、细胞周期的概念
细胞周期(cell cycle)是指从亲代细胞分裂结束到子代细胞分裂结束为止所经历的过程。
对于细胞周期的研究,一直是人们感兴趣的问题,随着研究的深入,对细胞周期的认识也逐渐深入。20世纪50年代前,人们把细胞的分裂过程划分为静止期(restingphase)和分裂期(Mphase)。直到1951年,Howard等采用放射自显影技术,用32P掺入洋葱根尖细胞以研究细胞内DNA的合成,首次提出细胞周期由G1、S、G2和M期组成的观点。进一步研究证明,此观点适用于大多数细胞类型。由于细胞化学、放射自显影和细胞分光光度术等新技术的应用,人们逐渐认识到细胞活动最活跃的时期是原来所谓的“静止期”(现在所说的间期),在此阶段完成DNA复制、RNA和蛋白质的合成,为细胞分裂提供物质准备。
细胞周期中,分裂期(M期)只占很短的时间,而绝大部分是细胞的生长期,即分裂间期(interphase)。间期中细胞要完成两个重要的生化活动:一是细胞质内的物质合成;二是细胞核内的DNA合成,这一很短的特定时段也称为DNA合成期,简称S期(DNAsynthesis)。研究表明,细胞在M期结束后,必须经历一段时间间隔才能进行DNA复制,M期结束到S期开始之间的间隙时间称为G1期(gap1),也称为DNA合成前期,是细胞生长、为DNA复制准备的阶段。此外,DNA复制完成后又必须经历一段时间才能进入M期,从S期结束到M期开始之间的间隙称为G2期(gap2),也称为DNA合成后期,主要为M期准备。M期细胞核的形态发生明显的变化,根据细胞核的变化特征将M期分为前期、中期、后期和末期。M期一结束,就形成两个子细胞(daughtercells),一个细胞周期即告结束,新生的子细胞又进入下一个周期(图8G2G1)。
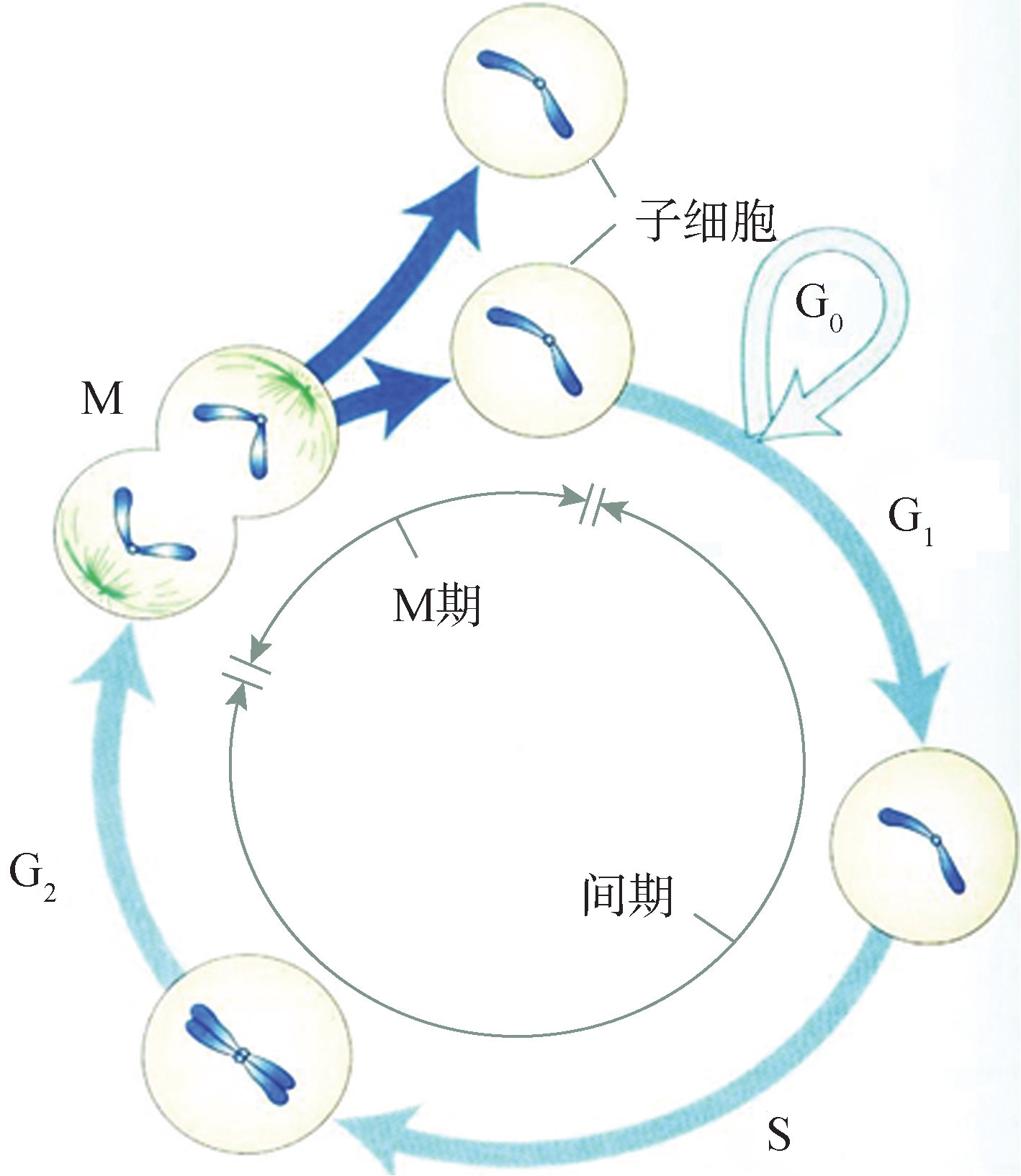
图 8G2G1 细胞周期示意
(转引自 LodishH 等, 2012
在正常的情况下,一个完整的细胞周期应包括4个时期,细胞沿着G1-S-G2-M期的路线运转。但在多细胞机体中,细胞的分裂行为有所差异。根据细胞的分裂行为,可将真核生物细胞分为3类:①持续分裂细胞,即细胞周期持续运转,又称为周期性细胞(cyclingcell),如小肠上皮细胞、皮肤基底层细胞、部分骨髓细胞等。②终末分化细胞,这类细胞一旦生成后,就不可逆地离开细胞周期,终生不再分裂,如哺乳动物的红细胞、神经细胞、肌细胞等。机体对终末分化细胞的需求依靠干细胞(stemcell)来补充。③暂不分裂细胞,又称为G0期细胞或休眠细胞,暂时从G1期离开细胞周期,但在给予适当刺激后可以重新进入细胞周期。如肝细胞、肾细胞及结缔组织中的成纤维细胞等。
二、细胞在周期各时相的变化
细胞分裂后即进入间期,在此期间要合成一系列物质为进入下一个分裂期做物质准备。实际上,G1期主要为S期做准备,而G2期主要为M期做准备。
(一)G1期的特征
G1期是细胞生长的主要阶段,在周期时间中所占的比例最大。此期合成细胞生长所需要的各种RNA、糖、脂等,RNA的合成导致结构蛋白和酶蛋白等形成,但不合成DNA,为进入DNA合成期做物质准备。实验证明,在G1期晚期到S期这一阶段,与DNA合成有关的酶和底物含量增高,尤其是DNA聚合酶的活性急剧升高(图8G2G2)。
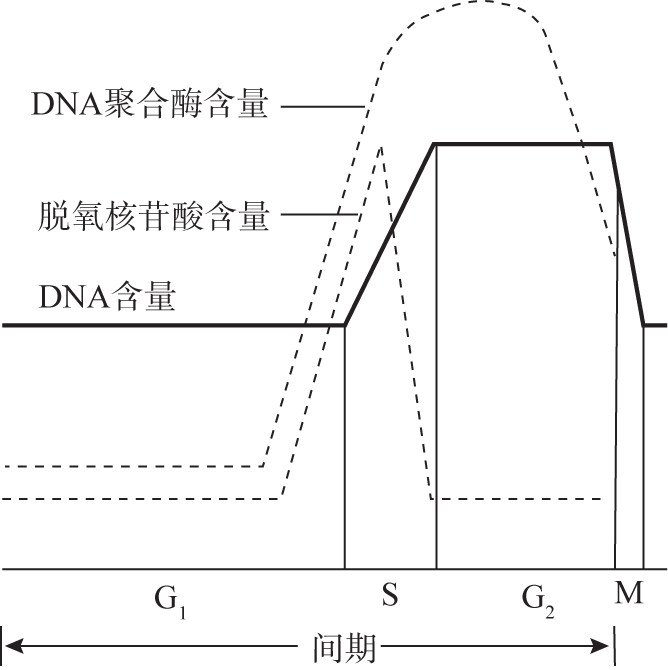
图 8G2G2 间期 DNA 合成的动态变化
关于G1期的调控早期研究主要集中在细胞周期的特性上。正常细胞从G1期晚期到S期阶段有1个或2个特殊的调节点,称为限制点(restrictionpoint,R点)或检验点(checkpoint)。R点起到了控制细胞增殖周期开和关的“阀门”作用。细胞是继续增殖还是进入静息(G0)期,是由它能否通过R点来决定的。当细胞处在不利条件下,如营养匮乏、药物等,细胞代谢速度降低,进入G0期以延长细胞生命;而肿瘤细胞往往失去全部或部分R点的控制,故细胞能不断地进行分裂。
(二)S期的特征
细胞进入S期后,开始进行DNA复制,同时参与组成核小体结构的组蛋白和非组蛋白等也在S期合成。DNA复制是细胞增殖的关键。DNA复制的起始和复制受到多种细胞周期调节因素的调控,如起始过程需要一种蛋白质性质的DNA合成诱导子的启动,即S期活化因子,这种因子由G1期过渡到S期开始合成,到S期中期含量最高,
S期结束时瞬间消失。另外,在S期还不断合成与DNA复制有关的酶,如DNA聚合酶、DNA连接酶等。新中心粒也在S期开始合成。
(三)G2期的特征
此期主要为M期准备物质条件,故G2期也称为丝裂前期(premitoticphase)。这一时期细胞主要合成与细胞分裂有关的蛋白质和RNA,如微管蛋白等,以及合成细胞进入M期促使染色质凝集的成熟促进因子(maturationpromotingfactor,MPF)。细胞通过G2期后能否顺利进入M期,受到G2期R点的控制,此期R点要检查DNA是否复制完成,细胞是否已生长到合适大小,环境因素是否有利于细胞分裂等。只有当所有利于细胞分裂的因素得到满足以后,细胞才能顺利由G2期过渡到M期。
(四)M期的特征
细胞周期中M期占用的时间最短,但细胞的形态结构变化最大。这一时期细胞的主要生化特点是RNA合成停止、蛋白质合成减少及染色体高度螺旋化。同时细胞形态学上发生极为显著的变化,这些形态上的变化,主要是保证将S期复制的DNA平均地分配到2个子细胞。丝状结构的产生、染色体的形成都是保证复制的两套遗传信息,在质和量上能够平均地分配给子细胞,以保证遗传的连续性和稳定性。M期中细胞核的分裂和细胞质的分裂在时间和空间上配合紧密,相互依赖、相互制约。
三、细胞周期的调控
对简单生物而言,细胞周期调控主要是为了适应自然环境,以便根据环境状况调节繁殖速度,以保证物种的繁衍。复杂生物的细胞则需要面对自然环境和其他细胞、组织的信号,并作出正确的应答,以保证组织、器官和个体的形成。细胞周期的精确调控对生物的生长、繁殖、遗传和变异都是十分重要的。这种高度的精确性一方面依赖于细胞内部的时钟调控,即周期蛋白依赖性激酶―周期蛋白(CDKsGcyclins)为中心的引擎周期变化所激发的一系列下游事件的发生,使细胞周期严格按照G1-S-G2-M期循环运转。另一方面在细胞周期异常时,细胞会采取补救措施进行调控,行使监控功能,避免差错的发生,如细胞周期的检验点(checkpoint)调控,可中断细胞周期使DNA进行修复。细胞周期调控与个体的生长、发育、衰老及细胞的癌变均密切相关。
(一)检验点在细胞周期中的作用
细胞周期中的多数时相均有限制点,如G1期、G2期和M期的检验点。这些检验点可调节细胞在细胞周期内按预定程序运行,如果细胞在未完成前一个时相内事件就进入后一个时相,就会出现遗传物质染色体复制不完全或染色体分配不均的错误。如用S期的细胞与M期的细胞融合,使S期细胞在未完成DNA复制的情况下提前进入M期,染色质提前凝聚成染色体,这种提前凝聚的染色体不是棒状的双体,而是碎裂的断片状,未完成DNA复制的情况下进入M期的细胞,其结果必然是死亡。而细胞从M期中期向后期进行的过程中,纺锤体微管与染色体动粒的连接时相也非常重要。如果在染色体动粒与微管尚未连接之前分裂后期就提前开始,就会造成分裂后的两个子细胞分配的染色体数目不等。如果这一过程发生在生殖细胞形成过程中,其后代就可以发生染色体异常疾病,其染色体核型发生错误。在正常情况下,细胞在细胞周期运行中发生这类错误的概率极小,其主要原因是由于检验点对细胞周期的监控(图8G2G3)。
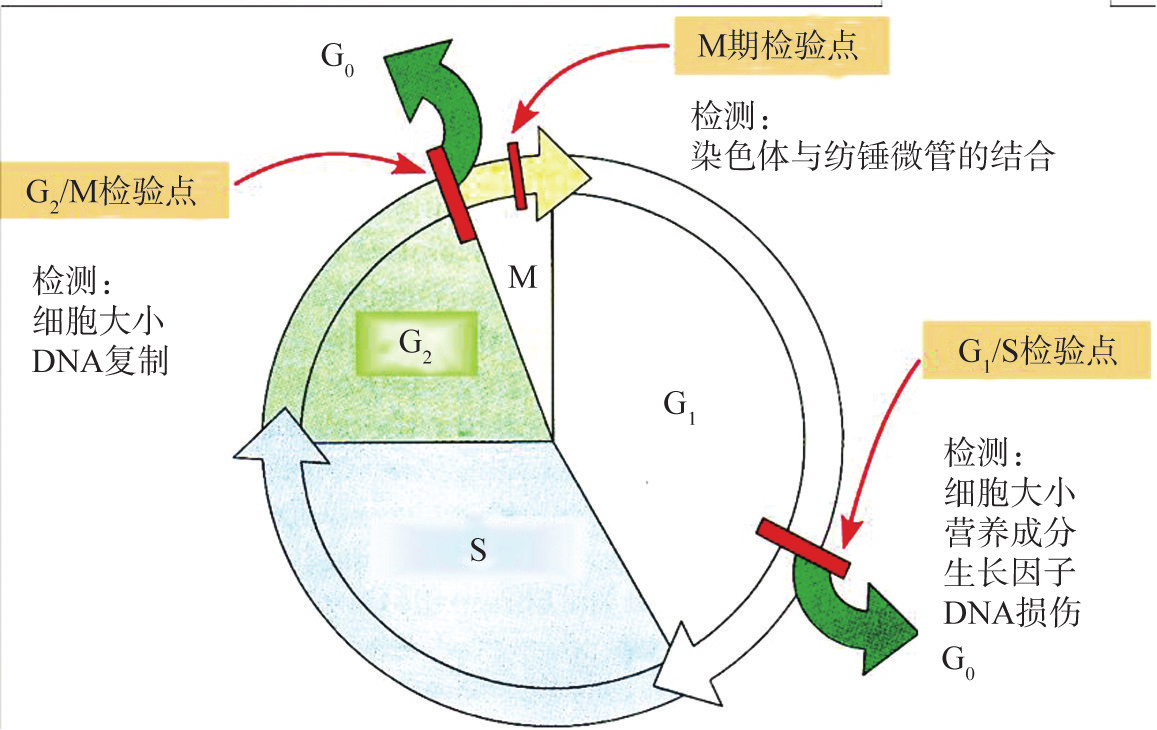
图 8G2G3 细胞周期检验点
在G1/S检验点,DNA损伤引起p53依赖的周期阻滞。正常细胞内p53的水平较低,DNA损伤刺激引起p53的表达和活性迅速升高。p53可诱导多种基因转录,如p21、Mdm2和Bax。p21是一种细胞周期抑制蛋白,通过抑制CDKs导致细胞周期阻滞,阻止损伤DNA的复制,使细胞周期停滞在G1期(图8G2G4)。Mdm2的作用是通过负反馈环调节p53蛋白水平,它可以结合并抑制p53的转录活性,有利于其通过泛素蛋白酶体途径降解。但细胞严重受损,损伤的DNA无法修复时,p53又可以激活某些基因的转录,如Bax、Fas及参与氧化应激反应的相关基因,诱导细胞凋亡。
G2/M检验点,当DNA损伤出现在G2期时,引起细胞周期阻滞,此作用可以不依赖于p53蛋白。细胞可以通过抑制CDK1的去磷酸化作用,使其处于抑制状态;或者通过将cyclinBlGCDKl复合物滞留在胞质中,使其不能进入细胞核发挥作用,故阻止细胞进入M期。
M期检验点,又称为纺锤体组装检验点,其作用是监测纺锤体形成过程中染色体不正确的组合,在有丝分裂中期引发周期阻滞,以阻止分裂后期启动和胞质分裂。几种哺乳动物M期检验点蛋白已被广泛研究,如Mad、Bub和Ndc80蛋白在微管黏附作用缺陷时被激活,其通过抑制后期促进复合物(anaphasepromotingcomplex,APC)的泛素连接酶活性,阻止分裂中后期的周期进展,避免染色体不正确地分离。
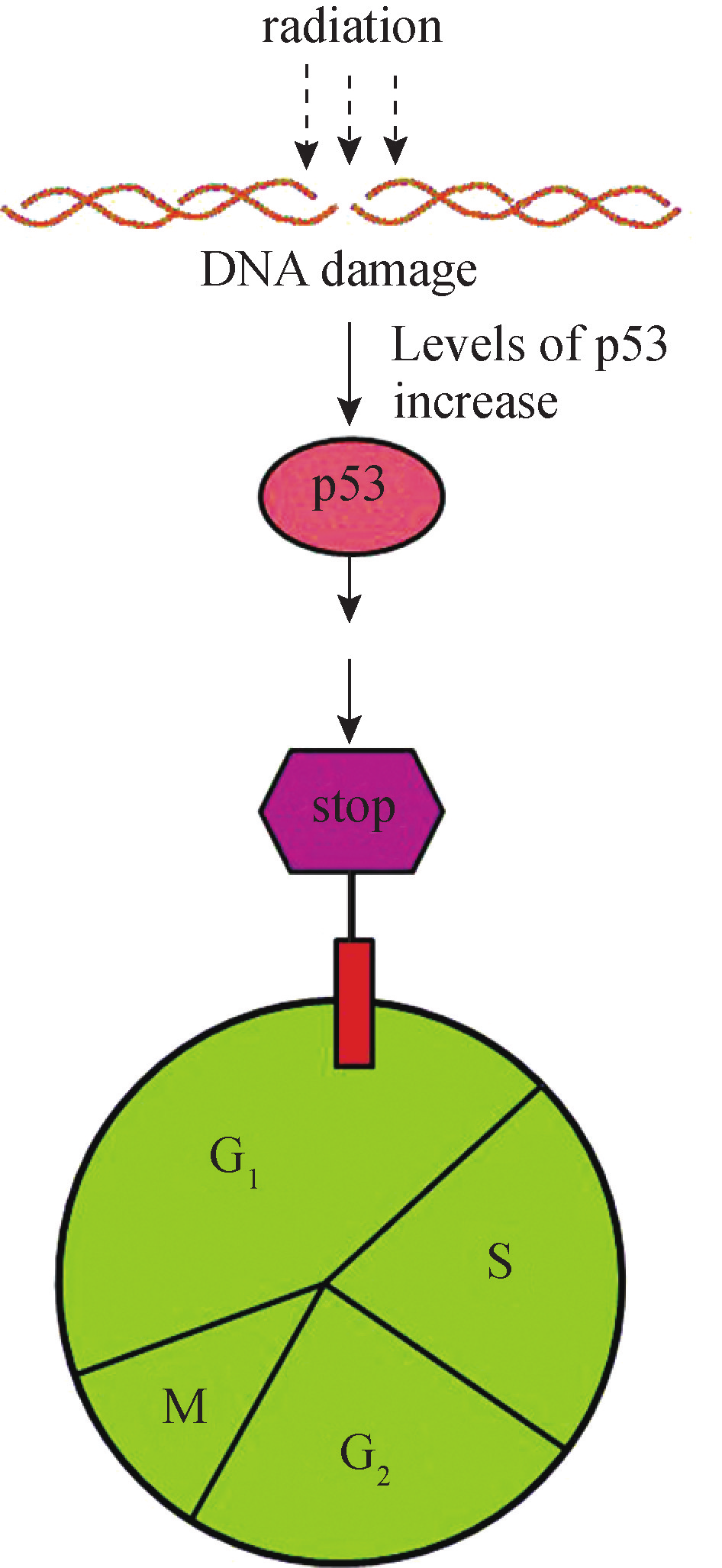
图 8G2G4 p53 在 DNA 损伤诱导 G 1 期阻滞中的作用
(二)CDK激酶和周期蛋白在细胞周期进程中的作用
在真核细胞周期逆转的调控研究中,发现与细胞分裂和细胞周期调控有关的基因,被称为细胞分裂周期基因(celldivisioncyclegene),简称cdc基因,该基因编码的蛋白质具有蛋白激酶活性,可以使各种蛋白质底物磷酸化,对一些生物的细胞分裂来说是必需的。在对细胞周期的深入研究中发现,cdc基因所表达的蛋白产物本身并不具有激酶活性,只有与周期蛋白(cyclin)结合后,其激酶活性才能表现出来,因而,它们被统称为周期蛋白依赖性蛋白激酶(cyclindependentkinase,CDK)。在CDK激酶复合物中cyclin为调节亚单位,CDK为其催化亚单位。迄今已发现多种cyclin,在哺乳类动物细胞中包括cyclinA、B、C、D、E、F、G、H等,它们分别与不同类型的CDK蛋白结合,构成不同的cyclin-CDK;不同的cyclin-CDK在不同的时相表现活性,影响不同的下游事件(表8G2G1)。
表 8G2G1 不同类型的 cyclins 和 CDKs
激酶复合体 | cyclin | CDK | 功能 |
| G 1CDKs 和 | cyclinD | CDK4和CDK6 | G1期 |
| G 1 / SCDKs | cyclinE | CDK2 | G 1 期至 S 期 |
| S CDKs | cyclinA | CDK2 | S 期 |
| M CDKs | cyclinB | CDK1 | M 期 |
G1期,在生长因子的刺激下,cyclinD表达,并与CDK4和CDK6结合,使下游的蛋白质如Rb磷酸化,Rb释放出转录因子E2F,促进多个基因的转录,如编码cyclinE、A和CDK1的基因。
在G1期至S期,cyclinE与CDK2结合,促进细胞通过R点进入S期。向细胞内注射cyclinE的抗体能使细胞停滞于G1期,说明细胞进入S期需要cyclinE的参与。S期的向前推进则需要cyclinA与CDK2形成的激酶复合物。同样将cyclinA的抗体注射到细胞内,发现能抑制细胞的DNA合成,推测cyclinA是DNA复制所必需的。在G
2期至M期,cyclinA、cyclinB与CDK1结合,CDK1使底物蛋白磷酸化,如将组蛋白H1磷酸化导致染色体凝缩,核纤层蛋白磷酸化使核膜解体等下游细胞周期事件。在M期当成熟促进因子MPF活性达到最高时,激活后期促进因子APC,通过泛素连接酶催化泛素与cyclin结合,cyclin随后被26S蛋白酶体降解,完成一个细胞周期(图8G2G5)。
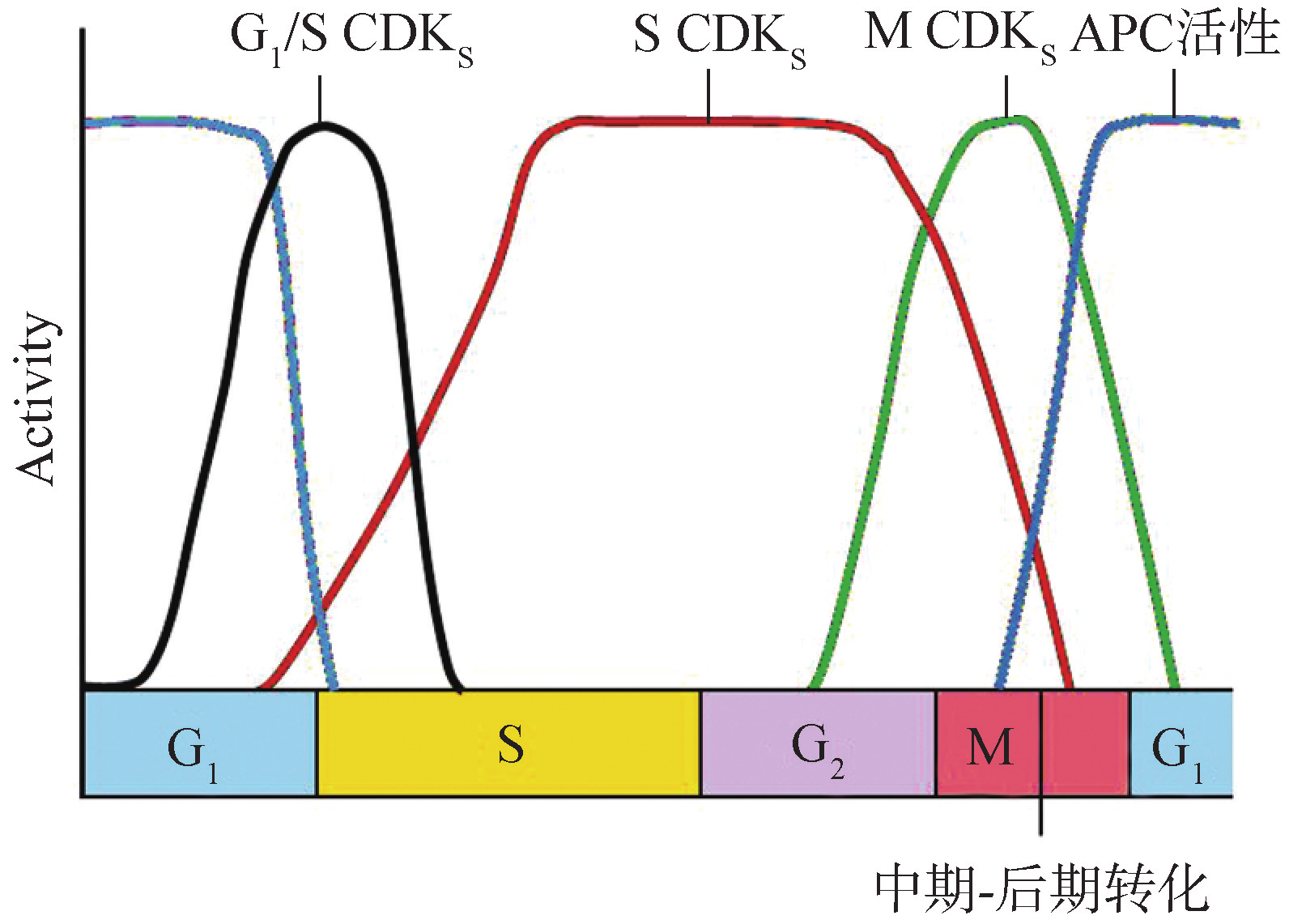
图 8G2G5 CDKs 调节细胞周期进程
细胞周期运转也存在负调控,研究表明,细胞至少可通过两种不同机制阻遏细胞周期的运转:一是周期调控系统组分停止合成,如G0期细胞中大部分Cyclin和CDKs都消失,这在多细胞生物尤其明显。二是CDKs的活性可以被CDK抑制物(CDKinGhibitor,CKI)所抑制,阻止cyclin-CDK复合物的装配或活性。现已发现两种CKI家族:
Cip/Kip家族和INK4家族。Cip/Kip家族包括p21(Waf1/Cip1)、p27(Cip2)
和p57(Kip2),可以广泛作用于cyclin-CDK复合物并抑制它们的活性,特别是G1期的cyclin-CDK4/6复合物。INK4家族包括p15(INK4b)、p16(INK4a)、p18(
INK4c)和p19(INK4d),它们均可以特异性抑制CDK4/6。CKI受胞内外的信号分子调节。例如,p21通过抑制增殖细胞核抗原(PCNA)而阻断DNA的合成。p15和p27的表达和激活可被转化生长因子TGF-β增强,通过多种途径抑制细胞周期进程。
因此,正常的细胞周期需要CDK的正调节因子cyclin与负调节因子CKI的精确协同与平衡,一旦这种平衡失稳就会造成细胞增殖失控。
(三)生长因子对细胞周期的影响
单细胞生物的增殖取决于营养是否足够,而多细胞生物的增殖取决于机体是否需要,这种需要是通过细胞通信来实现的。生长因子(growthfactor,GF)是一大类与细胞增殖有关的信号物质,目前发现的生长因子多达几十种,多数有促进细胞增殖的功能,故又称为有丝分裂原(mitogen),如表皮生长因子(EGF)、胰岛素样生长因子(IGF)、神经生长因子(NGF);个别具有双重调节作用,如转化生长因子β(TGF-β),能促进一类细胞的增殖,而抑制另一类细胞的增殖。
GF没有种属特异性,但有很强的组织特异性,即不同种类的细胞需要不同的GF。现已分离出几十种GF,它们普遍存在于机体的各种组织中(表8G2G2)。GF受体也普遍存在,许多细胞表面同时存在一种以上的GF受体,能接受不同GF的顺序性调节,即所谓GF的协同作用。例如,处于G0期的3T3细胞必须经过血小板衍生生长因子(PDGF)的激活才能进入G1期;G1期进入S期又要经过EGF和IGF的顺序激活。
为了防止机体细胞过度生长,除具有细胞增殖的正向调节因子外,还具要有负调节因子,以平衡细胞的增殖活性。抑素是一类细胞中产生的可以抑制细胞增殖的调节因子,有些是小分子可溶性蛋白,有些是糖蛋白。抑素没有种属特异性,但有严格的组织和细胞特异性,只对同类细胞具有抑制作用。例如,上皮抑素可以抑制上皮细胞的增殖;肝抑素抑制肝细胞的增殖。
目前发现与生长因子有关的信号通路主要有:EGF途径、TGF-β途径、cAMP途径和磷酯酰肌醇途径。例如,EGF与受体(EGFR)结合后,可以使受体发生二聚体化,从而改变受体的构象,使其中的酪氨酸激酶活性增强,进一步活化Ras,激活的Ras作用于MAPK激活系统,导致ERK的激活,最后ERK转位至细胞核内,促进细胞增殖相关基因的表达。如通过一种未知的途径激活c-myc,myc作为转录因子促进cyclinD、E2F等许多G1-S相关基因的表达,细胞进入G1期。
表 8G2G2 几种生长因子的特性
| 生长因子 | 组成 (分子量) | 组织来源 | 靶细胞 |
表 皮 生 长 因 子 ( EGF ) | 含 53 个氨基酸的多肽 (6000 ) | 颌下腺 | 上皮细胞等多种细胞 |
胰岛素样生长因 子 (IGF ) | 67/ 70 个氨基酸残基组成的多 肽 (8000 ) | 肝 细 胞 合 成,存 在于血浆中 | 骨组织等多种细胞 |
血小板衍生生长 因子 ( PDGF ) | 二聚体蛋白 ( 30000 ) | 巨 细 胞 合 成,存 在于血小板中 | 成 纤 维 细 胞、胶 质 细胞、平滑肌细胞等 |
生长调节素 C ( SM-C ) | 70个氨基酸残基组成的多肽( 7000 ) | 肝 细 胞 合 成,存 在于血浆中 | 骨组织等多种细胞 |
(四) 与细胞周期有关的基因
(四)与细胞周期有关的基因
1。细胞分裂周期基因与细胞周期 在细胞内有一类与细胞周期运转和调控有关的基因,称为细胞分裂周期基因(cdc)。细胞周期运行的有序性与cdc基因在细胞周期的不同阶段表达有关。例如,在酿酒酵母(S。cerevisiae)中已确定了一些cdc基因的表达顺序及可能的生物学功能,在细胞周期起始阶段主要受cdc28调控;cdc8作用于DNA合成起始;纺锤极体(SPB)的复制被cdc31控制;cdc24控制酵母的出芽(图8G2G6)。
实验证明,裂殖酵母(S。pombe)的cdc2基因与酿酒酵母的cdc28基因具有同源性,哺乳动物(包括人)与cdc2同源的基因产物是分子量34000的蛋白质p34,其基因序列与cdc2/cdc28之间的同源性是63%。
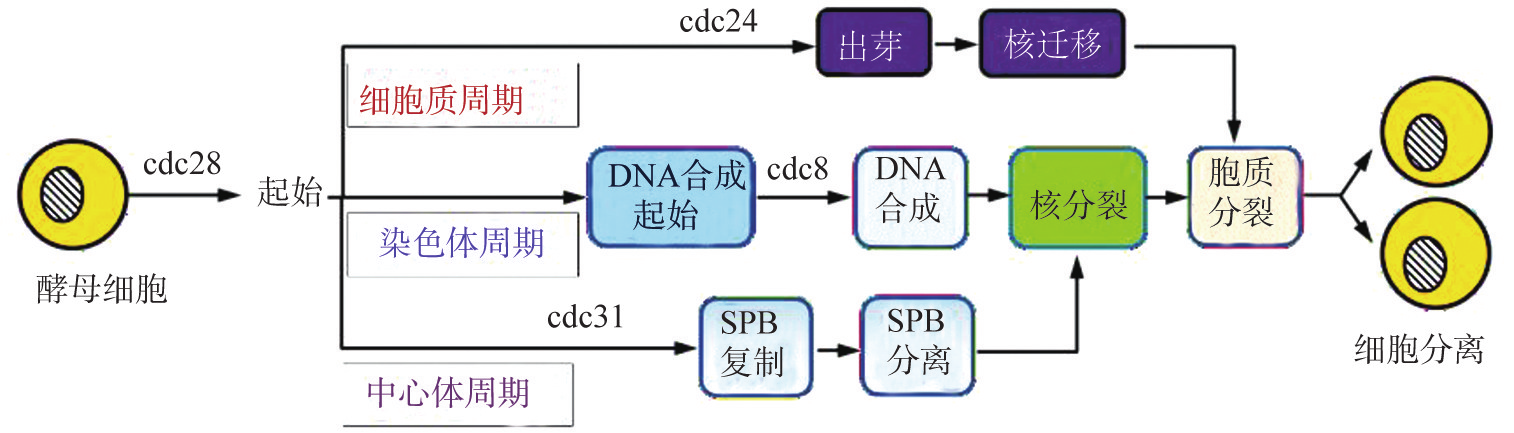
图 8G2G6 酿酒酵母周期中 cdc 基因的调控作用
癌基因和抑癌基因与细胞周期 在正常细胞的基因组中,含有与病毒癌基因 ( v-oncogene )相似的原癌基因 ( proto-oncogene ).原癌基因编码的蛋白质产物包括生长因子、生长因子受体、细胞内信号分子等,都与调节细胞的增殖和分化活动有关.原癌基因的产物是正常细胞增殖所必不可少的,但是它们一旦发生突变成为癌基因 (onco-gene ),就 会 引 起 正 常 细 胞 无 限 增 殖 最 终 导 致 肿 瘤 形 成. 而 抑 癌 基 因 (tumor suppressorgene )实际上是正常细胞增殖过程中的负调控因子,它编码的蛋白质在细胞周期的检验点上起着阻止周期进程的作用.如果抑癌基因突变,丧失其细胞增殖的负调控作用,则导致细胞周期失调而过度增殖.
Rb基因和p53基因是两类与人类恶性肿瘤关系最为密切的抑癌基因。Cyclin、CDK、CKI对细胞周期的调控主要通过Rb途径、p53途径来实现。Rb是从遗传性视网膜母细胞瘤中发现的第一个抑癌基因,主要在G1/S检验点发挥作用。在G1期,Rb蛋白被cyclinGCDK复合物磷酸化后,释放转录活化因子E2F,使E2F的应答基因转录,并使细胞进入S期(图8G2G7)。E2F和cyclin之间可能有一个潜在的正反馈调节环路。p53的调控机制是阻止细胞周期停滞在G1期,为细胞提供更多的时间修复损伤的DNA。
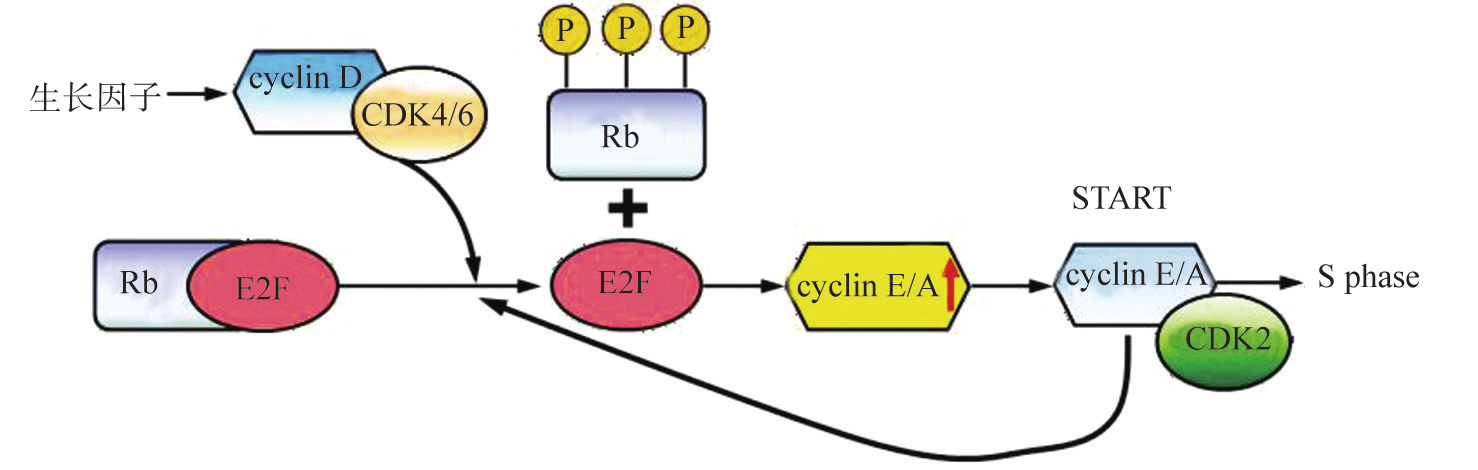
图 8G2G7 Rb 调控 G 1 / S 期转化

