
第一节 细胞分裂
细胞的分裂增殖是生命得以延续的保证。细胞分裂主要有三种方式:无丝分裂、有丝分裂和减数分裂。
一、无丝分裂
无丝分裂(amitosis)是最早被发现的一种细胞分裂方式,是指处于间期的细胞核不经过任何有丝分裂时期,而分裂为大小大致相等的两部分的细胞分裂方式。早在1841年雷马克(Remak)在鸡胚的血细胞中观察到。1882年,弗来明(Flemming)发现其分裂过程有别于有丝分裂,因为分裂时没有纺锤丝与染色体的变化,所以称为无丝分裂。又因为这种分裂方式是细胞核和细胞质的直接分裂,所以又称为直接分裂。
无丝分裂在低等植物中普遍存在,在高等植物中也常见。高等植物营养丰富的部位,如胚乳细胞、表皮细胞、根冠等,无丝分裂很普遍。人体大多数腺体都有部分细胞进行无丝分裂,主要见于高度分化的细胞,如肝细胞、肾小管上皮细胞、肾上腺皮质细胞等;蛙的红细胞、蚕的睾丸上皮细胞均进行无丝分裂。无丝分裂周期包括4个时期(图8G1G1):

图 8G1G1 植物细胞的无丝分裂
第1期:核内染色质复制倍增,核及核仁体积增大,核仁及核仁组织中心分裂。
第2期:以核仁及核仁组织中心为分裂制动中心,以核仁与核膜周染色质相联系的染色质丝为牵引带,分别牵引着新复制的染色质和原有的染色质。新复制的染色质在对侧核仁组织中心发出的染色质丝的牵引下,离开核膜移动到核的赤道面上。
第3期:为核拉长呈哑铃形,中央部分缢缩变细,这是因为赤道面部位的核膜周染色质不与核膜分离,而核仁组织中心发出的染色质丝(与核膜周染色质相联系)螺旋化加强,产生的牵引拉力导致赤道面部位的核膜内陷。
第4期:核膜内陷加深,缢裂成两个完整的子细胞核,细胞随之分裂成两部分。由于无丝分裂的过程不出现染色体和纺锤丝,分裂后遗传物质不一定平均分配给两个子细胞。
二、有丝分裂
有丝分裂(mitosis)是大多数生物体细胞增殖的主要方式。在有丝分裂过程中,遗传物质复制一次,细胞分裂一次,细胞的遗传物质均等分配到两个子细胞中,保证了遗传物质在传递过程中的相对稳定性。
在细胞分裂过程中,细胞核的分裂和细胞质的分裂在时间和空间上紧密配合,相互依赖,相互制约,构成一个复杂而连续的动态过程。根据细胞分裂期核膜、染色体、纺锤体装配及核仁等形态结构的规律性变化,将有丝分裂过程分为前期、中期、后期和末期(图8G1G2)。
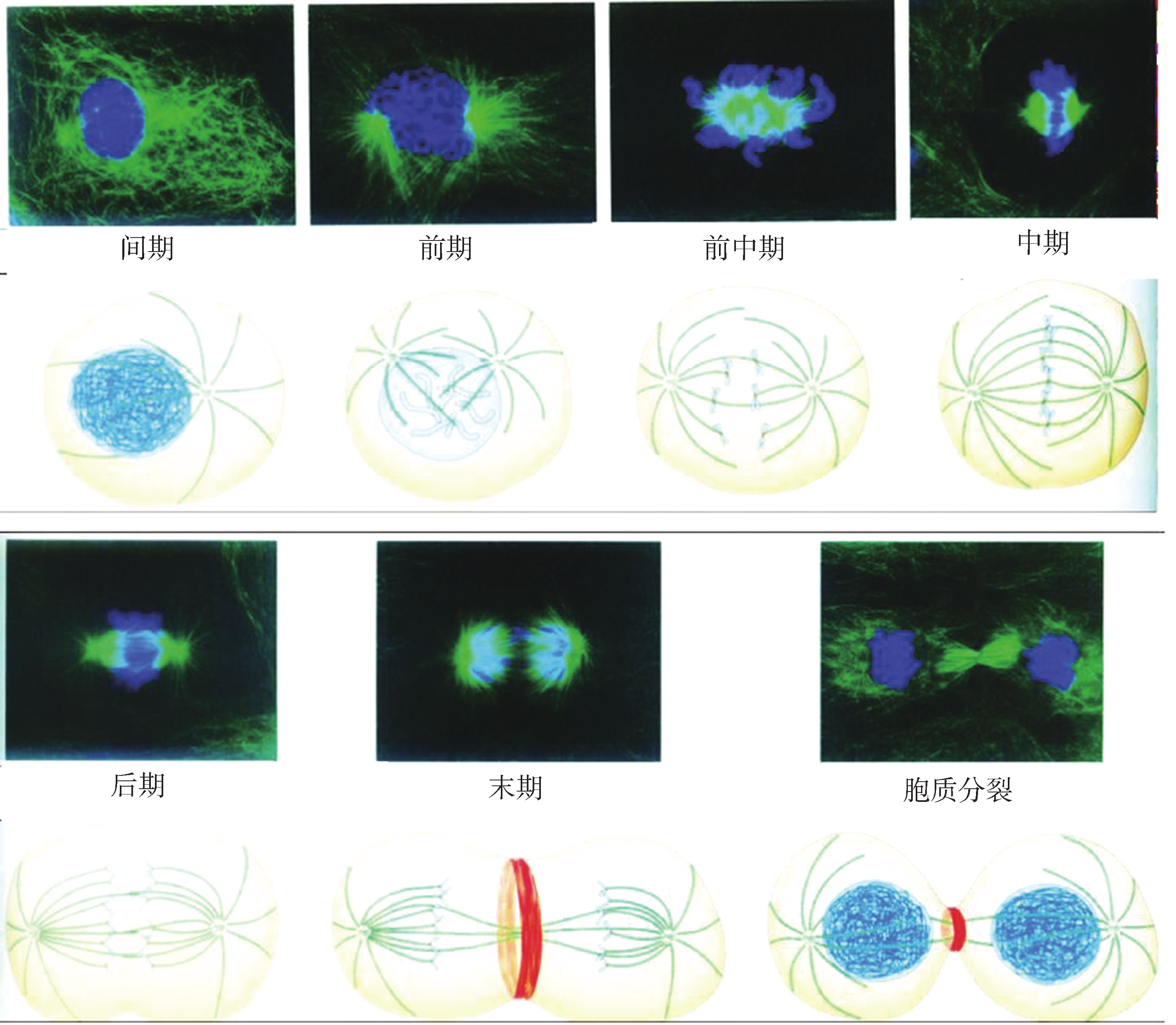
图 8G1G2 动物细胞的有丝分裂
(转引自 LodishH 等, 2012
1 前期 前期 (prophase)发生的主要变化包括:染色质凝集、核仁解体、核膜破裂。染色质纤维经螺旋化、折叠,最终形成棒状或杆状的染色体,每条染色体由两条染色单体构成。
2中期 中期 ( metaphase )染色体达到最大程度的凝缩,着丝粒两侧的动粒分别与本侧的动粒微管相连。在微管牵引下,染色体向赤道面集中形成赤道板,呈现出最具典型特征的中期染色体形态结构。赤道板平面与纺锤体纵轴垂直。采用秋水仙素等抑制微管聚合的药物处理细胞,可阻断细胞分裂,使其停滞在中期,从而获得大量的中期分裂相。出现典型的有丝分裂器,参与随后发生的染色体分离、染色体向两极移动及平均分配到子细胞等过程。
3后期 后期(anaphase)时,每条染色体的着丝粒纵裂,细胞内的染色体被平均分成两组,在纺锤体微管牵引下,两组染色体分别向细胞的两极移动。后期染色体向两极运动的动力来自动粒微管的不断缩短和极间微管的不断伸长。后期结束时,形态和数目相同的两组染色体到达细胞两极。
4末期 末期 ( telophase )的主要变化是子细胞核形成和胞质分裂。到达两极的染色体解螺旋重新转变为染色质;消失的核仁在核仁组织区 (nucleolarorganizingreGgion , NOR )的位置上开始重新出现;分散在胞质中的核膜小泡与染色质表面相连,并相互融合,形成核膜。至此,两个子细胞核形成。胞质分裂开始时,赤道面周围的细胞膜内陷,其下方肌动蛋白和肌球蛋白大量聚集,形成收缩坏 (contractilering ),继而形成分裂沟,后者不断加深,使细胞一分为二,完成胞质分裂。
三、减数分裂
减数分裂(meiosis)是有性生殖个体的配子形成过程中的一种特殊分裂方式,又称作为熟分裂(maturationdivision)。减数分裂的主要特点是:DNA复制1次,细胞连续分裂两次,子细胞中遗传物质减半。构成减数分裂的两次分裂称为减数分裂Ⅰ(meiosisⅠ)和减数分裂Ⅱ(meiosisⅡ)(图8G1G3),之前还包括减数分裂前间期。
(一)减数分裂过程
1. 减数分裂前间期 ( preGmeiosisinterphase ) 是为减数分裂做准备的阶段,需要经过较长的生长过程,进行足够的物质积累,最后形成初级生殖母细胞.该阶段也分为 G1、 S 和 G 2 期,与有丝分裂间期相比有3方面不同: ①S 期明显延长; ② 染色体只在一侧有动粒,所以在第 1 次减数分裂时姐妹染色单体不分离,共同进入一个子细胞;③G2期具有细胞增殖的限制点 ( R 点),有些种类的生物及人的卵母细胞长期停滞在G2期,只有接受性激素刺激后才能进行第 次减数分裂。
2.第1次减数分裂 减数分裂I的分裂时间比减数分裂Ⅱ的分裂时间长,在此时间内,同源染色体配对和遗传物质交换重组为其主要特征.进一步可细分为前期 I (包括细线期、偶线期、粗线期、双线期和终变期)、中期 I 、后期 I 和末期 I .
(1)前期I。此期的时间非常长,变化也非常复杂。根据细胞核的形态变化可划分为细线期(
leptotenestage)、偶线期(zygotenestage)、粗线期(pachytenestage)、双线期(diplotenestage)和终变期(diakinesisstage)。
1)细线期:染色体呈细丝状,相互交织成网状。染色质丝开始凝缩。在此期之前DNA已经复制,每条染色质丝中含有2条染色单体,但是光学显微镜下仍呈细线状,在细线上可见深染的、由染色质丝盘曲而成的染色粒,染色体端部开始与核膜附着斑相连,有利于同源染色体配对。
2)偶线期:最典型的特征是染色体配对。随机分布的同源染色体通过其一侧形成侧体彼此识别并相互配对,配对后两个侧体之间形成轴体结构,像拉链一样使同源染色体紧密相连。同源染色体配对的过程称为联会(synapsis),侧体与轴体总称为联会复合体(synaptonemalcomplex,SC),其化学成分主要为蛋白质。联会的结果是每对同源染色体形成一个密切相伴的二价体(bivalent)。
3)粗线期:染色体进一步螺旋化,变短变粗,在光学显微镜下可以看到每条染色包含2条染色单体,互称姐妹染色单体(sisterchromatid)。每个二价体都由2条同源染色体组成。这样一个二价体有4条染色单体,称为四分体(tetrad)。同源染色体的染色单体之间互称为非姐妹染色单体(non-sisterchromatid)。在此期间,发生同源染色单体的横向断裂,并在断裂处发生同源染色体非姐妹染色单体之间的交换。
4)双线期:染色体进一步螺旋化而缩短。同源染色体之间的联会复合体解体。同源染色体相互排斥趋向分离,使互换后的染色体出现交叉(chiasma)。一般认为,交叉是同源染色体的非姐妹染色单体交换的形式。
5)终变期:染色体更加变粗变短,并向核的四周移动,纺锤体开始形成,核仁、核膜消失。交叉移行到染色体的末端,交叉数量减少。
(2)中期I。各二价体排列在赤道面上形成赤道板,纺锤体微管与染色体一侧的动粒相连,两条同源染色体的动粒各朝向细胞的一极,但二价体仍存在少数交叉相连。
(3)后期I。同源染色体之间彼此分离,随机分配给2个子细胞。每个子细胞只获
得了1对同源染色体中的1条,即二分体(dyad)。
(4)末期I。二分体到达细胞两极并逐渐解旋成为染色质,核膜、核仁重现;胞质分裂,形成两个子细胞。此时,每条染色体虽然有2条染色单体,但染色体数目减少了一半,为单倍体(n)。在人类,由减数分裂产生的子细胞染色体数目是23。
3. 减数分裂间期 完成第1次减数分裂后,细胞进入短暂的分裂间期。此期没有新的 DNA 合成,只进行动粒的组装和中心粒复制.间期持续的时间随不同的生物种类差异较大。
4. 第2次减数分裂 减数分裂Ⅱ的过程和有丝分裂基本相同。前期有纺锤体形成,动粒微管连接到染色体的动粒上,牵引染色体运动。染色体排列形成赤道板,细胞即进入短暂的中期.随后着丝粒断裂,姐妹染色单体彼此分离,分别向纺锤体的两极移动,最终细胞发生一分为二等后期和末期事件。经过上述的两次减数分裂,由 1个母细胞分裂成 4个子细胞。子细胞的染色体数目只有母细胞的1/2,成为单倍体的生殖细胞。
动粒微管连接到染色体的动粒上,牵引染色体运动。染色体排列形成赤道板,细胞即进入短暂的中期。随后着丝粒断裂,姐妹染色单体彼此分离,分别向纺锤体的两极移动,最终细胞发生一分为二等后期和末期事件。经过上述的两次减数分裂,由1个母细胞分裂成4个子细胞。子细胞的染色体数目只有母细胞的1/2,成为单倍体的生殖细胞。
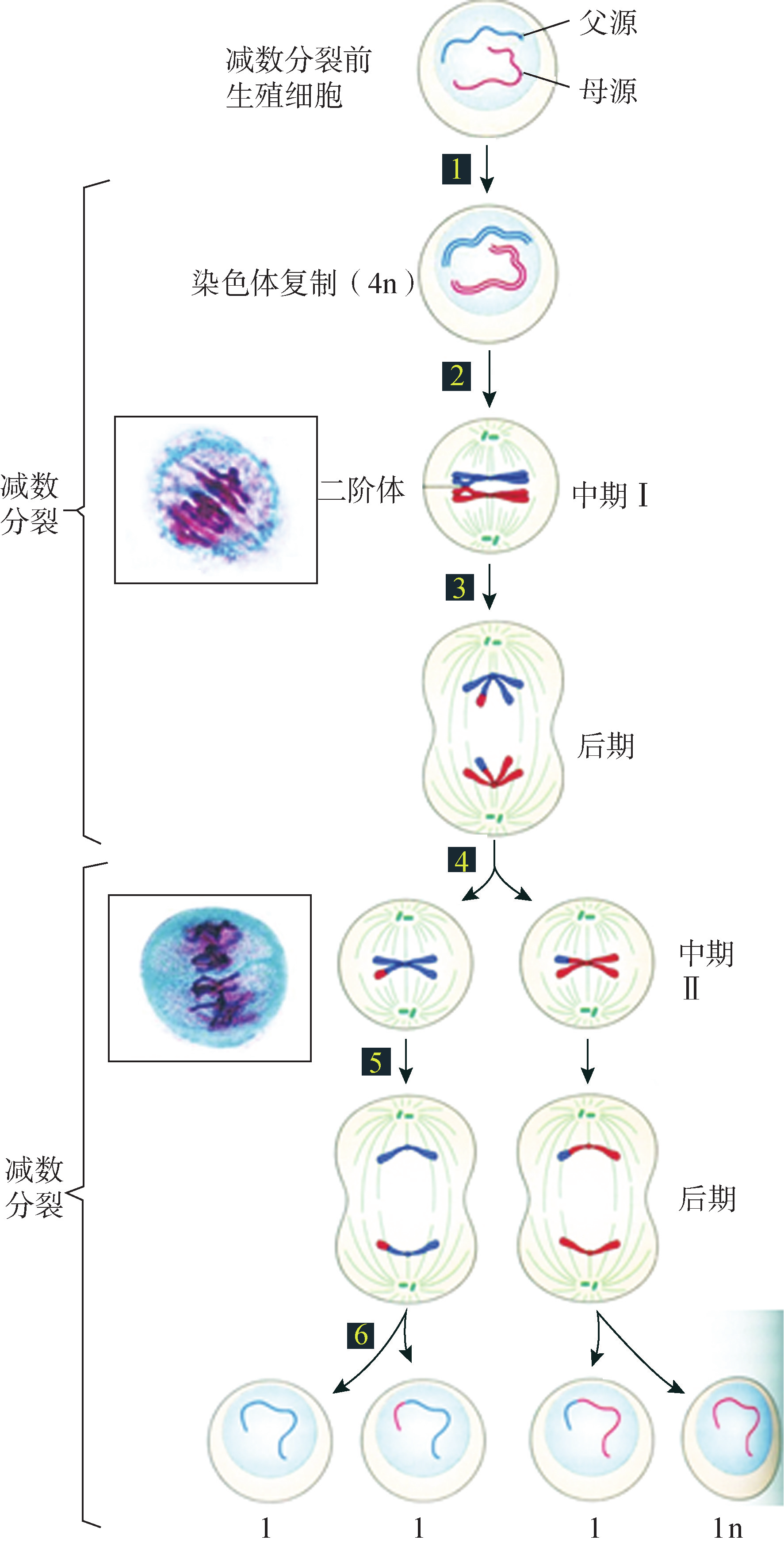
图 8G1G3 减数分裂基本过程
(转引自 LodishH 等, 2012 )
(二)减数分裂与有丝分裂比较
在减数分裂和有丝分裂过程有许多相同之处。例如,都有细胞核与细胞质的周期性变化,都形成有丝分裂器,都存在染色体的螺旋化和去螺旋化等。但两者之间也存在明显区别。
减数分裂过程中DNA复制1次,细胞连续分裂两次,1个亲代细胞形成4个遗传组成不同的子细胞,且遗传物质只有亲代细胞的一半,产生遗传多样性。在有丝分裂过程中,DNA复制1次,细胞分裂1次,1个亲代细胞形成两个遗传组成完全相同的子细胞,保证了遗传的稳定性。
减数分裂是配子形成特有的细胞分裂,减数分裂I中发生同源染色体的配对及非姐妹染色单体之间的交换;有丝分裂则是体细胞的增殖方式,细胞中每条染色体都是独立的,不产生联会和交换。
(三)配子发生
有性生殖生物的精子与卵子的形成过程称为配子发生(gametogenesis)。在形成过程中,精子和卵子虽然是来自不同性别的亲代,经历的过程也有一定的差异,但却有一个共同的特点,即经过一系列的有丝分裂后,在成熟期都要进行减数分裂。人类精子和卵子的形成过程均需要经历增殖期、生长期、成熟期,精子细胞的产生还要经过变形期(图8G1G4)。1个精母细胞经过2次减数分裂,产生4个均一的、具有生理功能的精子;1个卵母细胞经第1次减数分裂形成1个很大的次级卵母细胞和1个很小的第1极体(只有细胞核)。第2次减数分裂又产生1个很大的卵细胞和1个很小的第2极体,同时第一极体也一分为二。所以一个卵母细胞经过2次减数分裂产生1个卵细胞和3个极体,它们都是单倍体(n=23),3个第二极体均退化。
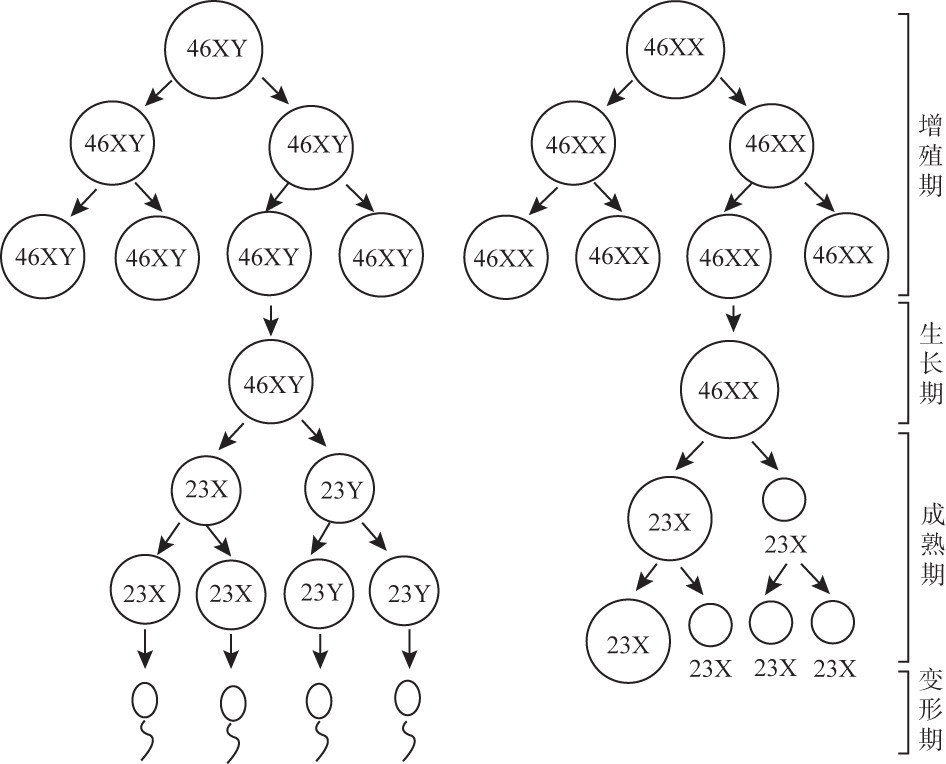
图 8G1G4 精子与卵子的发生过程

