
第一节 核 膜
学习要求
知识内容
核膜(nuclearmembrane)又称核被膜(nuclearenvelope),是位于真核生物细胞分裂间期细胞核最外面的不对称的双层膜结构。核膜是细胞内膜系统的一部分,它的出现使细胞核成为细胞中一个相对独立的体系。
一、核膜的化学组成和结构
(一)核膜的化学组成
核膜的化学成分主要为蛋白质和脂类,这与内质网很相似。核膜中蛋白质占65%~75%,脂类(多为磷脂)占25%~40%,糖类约占5%,另外,可能有微量的DNA、RNA及金属离子等。其中含有20多种蛋白质,分子量为16000~160000,包括组蛋白、基因调节蛋白、核纤层蛋白、与电子传递有关的酶、DNA和RNA聚合酶、以及RNA酶等。定位在核膜不同部位的蛋白质分别行使不同的功能。核膜中含有的脂类包括不饱和脂肪酸、磷脂酰乙醇胺、胆固醇和三酰甘油等。核膜与内质网在组成成分上具有相似性和特异性,如核膜含有较低含量的不饱和脂肪酸和较高含量的胆固醇和三酰甘油,这表明核膜与内质网既具有密切关系,又拥有各自的结构特点。
(二)核膜的形态结构
核膜主要由两层平行且不连续的非对称性单位膜构成,每层单位膜厚度约为7.5nm,包括外核膜、内核膜、核周间隙、核纤层和核孔复合体等结构(图7G1G1)。
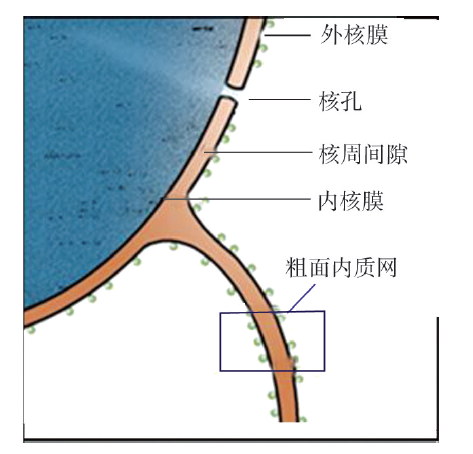
1. 外核膜(outernuclearmembrane)
外核膜是核膜中面向细胞质的一层单位膜,在形态及生化行为上与粗面内质网近似,表面有核糖体附着,可进行蛋白质的合成,并与粗面内质网相连,使核周隙与内质网腔相通。因此,外核膜被认为是粗面内质网的特化区域。在外核膜的胞质面附着有微管和中间纤维形成的细胞骨架成分,与细胞核在细胞内的定位及维持细胞核的形态有关。
2.内核膜(innernuclearmembrane)内核膜是核膜中面向核基质的一层单位膜,与外核膜平行排列,表面光滑无核糖体附着,内表面附着核纤层。内核膜上有特异蛋白,如核纤层蛋白B受体为核纤层蛋白B提供结合位点,从而将核膜固定在核纤层上,起到固定核膜,稳定核形态的作用。
3. 核周间隙(perinuclearspace)核周间隙也称核周池,是位于内外核膜之间的腔隙。宽20~40nm,这一宽度常随细胞种类不同而不同,在同一细胞核的不同部分也不完全一致,且随细胞的功能状态而改变。它与粗面内质网腔相通,含有多种蛋白质、酶及离子等。核周间隙不仅是胞质和核质间进行物质交换的重要通道,而且也是核与质间的生理缓冲区域。
4. 核纤层(nuclearlamina)核纤层是内核膜内侧与染色质间的一层电子密度较高的纤维状的蛋白网格结构。它与中间纤维和核骨架相互联系,形成贯穿于细胞核和细胞质的骨架体系。其厚度随细胞不同而不同,多数为10~20nm,纤维直径约为10nm。核纤层与内核膜紧密结合,整体呈球状或笼状网络,切面呈片层结构(图7G1G2)。

图7G1G2 核纤层的超微结构
核纤层的主要成分是核纤层蛋白(lamin)。哺乳动物和鸟类细胞的核纤层蛋白含有3种属于中间纤维性质的多肽,分别为LaminA、LaminB、LaminC,其相对分子量为60~75kD。研究证明,LaminA与LaminC是同一基因编码的不同加工产物,三种类型的核纤层蛋白均以二聚体的形式存在,有杆状的α螺旋中心和球形的头部及尾部。众多的核纤层蛋白单体组装形成核纤层。在真核生物的间期细胞核中,LaminA和LaminC以头-头、尾-尾相接的方式形成核纤层纤维,并通过LaminB固着于内核膜表面;在分裂期,核纤层解体,LaminA和LaminC以蛋白单体的形式存在于细胞质中。
核纤层与核膜、染色质及核孔复合体在结构上紧密联系,推测其可为核膜及染色质提供结构支架,稳定细胞核的形态结构,参与染色质的凝集、DNA的复制、RNA的转录及核膜的崩解与重建。
5. 核孔复合体(nuclearporecomplex,NPC)
核孔复合体是内外核膜在某些部位融合形成的由核孔蛋白组成的蛋白质复合物环状小孔。它是细胞核与细胞质之间进行物质运输的重要通道。孔径为80~120nm,大多数为50~70nm。一个典型的哺乳动物细胞核膜上含有300~4000个核孔,约占膜面积的8%。核孔的数目、疏密程度及分布形式与细胞类型和生理状况有关。在分化程度较低、合成代谢旺盛和核仁大的细胞中,核孔数目较多,反之较少。一般来讲,动物细胞的核孔数目要多于植物细胞。如在高度分化但代谢活跃的肝、肾等细胞中核孔数为12~20个/μm2;在代谢低或增殖不活跃的红细胞中核孔数仅为1~3个/μm2;而许多成熟精子的核膜上几乎看不到核孔。
核孔复合体主要由蛋白质组成,估计有50~100多种蛋白质。在酵母中已鉴定出30多种核孔蛋白。迄今已鉴定的脊椎动物核孔复合体蛋白成分也达10多种。核孔复合体在进化上高度保守。免疫交叉反应表明,从酵母到人的核孔蛋白之间具有很高的同源性,且核孔复合体的整个结构在进化上也具有很高的保守性。
核孔复合体一直以来是细胞形态学研究的目标之一,具有多种模型,其中被普遍接受的是捕鱼笼式(fishGtrap)模型(图7G13),其内径为80~120nm,外径为120~150nm,具体结构包含胞质环、核质环、中央栓和辐四部分。

图7G1G3 核孔复合体结构模型(摘自BAlberts)
(1)胞质环(cytoplasmicring):又称为外环,是位于核孔边缘胞质面一侧的环状结构,与柱状亚单位相连,其上有8条短而弯曲的胞质纤维,对称分布,伸向细胞质。(2)核质环(nucleoplasmicring):又称为内环,是位于核孔边缘核质面一侧的环状结构,与柱状亚单位相连,其上有8条细长纤维,对称分布,伸向核质,在纤维末端又形成一个由8个颗粒组成的小环,这样就形成一个捕鱼笼似的结构,称为核篮(nuclearbasket)。
(3) 中央栓(centralplug):又称为中央颗粒,位于核孔复合体中心,呈颗粒状或棒状,其实质是中央运输蛋白,在核质交换中发挥一定作用。
(4) (4)辐(spoke):是由核孔边缘伸向中央栓的8个辐条结构,呈辐射状八重对称,将胞质环、核质环、中央栓连为一体。辐的外侧有外辐条环,内侧有内辐条环,进一步可分为3个结构域:①柱状亚单位(columnsubunit):是辐的主要区域,位于核孔边缘,连接胞质环与核质环,可以起到支撑作用;②腔内亚单位(luminalsubunit):是位于柱状亚单位外侧,穿过核膜伸入双层核膜的核周间隙的区域,可以起到锚定作用;③环状亚单位(annularsubunit):是位于柱状亚单位内侧,由粒状结构环绕形成的区域,靠近核孔复合体中心部分,与内辐条环相连,形成核质交换的通道。
核孔复合体的转运物质方式有被动运输和主动运输两种,可介导生物大分子的入核和出核的双向转运,并对蛋白质等生物大分子物质的转运具有选择性。在特定信号引导下,核孔复合体可将构成核糖体、染色质的蛋白质及细胞核代谢所需的各种酶运入细胞核,也可将在细胞核内合成的各种RNA和装配形成的核糖体亚基等物质运输到细胞质中,完成细胞核与细胞质之间的大分子物质的交换。
二、核膜的功能
二、核膜的功能
(一)核膜的区域化作用
核膜的区域化作用使DNA的复制、RNA的转录与蛋白质的翻译在空间上分离(图7G1G4)。核膜构成细胞核与细胞质间的一道天然选择性屏障,将细胞分为两大结构区和功能区。原核细胞无核膜分隔,其RNA的转录与蛋白质的合成在同一空间进行,RNA 分子未完成转录就开始与核糖体结合并指导蛋白质的合成。而真核细胞的DNA复制、RNA 的转录与加工在细胞核内进行,RNA 前体在细胞核内先进行有效的剪切,去除内含子,并进行必要的修饰,加工成熟后的mRNA 才能进入细胞质指导蛋白质的合成。核膜的出现使细胞核具有一个相对稳定的内环境,使遗传信息的传递在不同的空间进行,避免彼此的干扰,确保基因表达的准确性与高效性。

图7G1G4 核膜的区域化作用
(二) 控制细胞核与细胞质之间的物质交换
细胞核与细胞质之间的物质交换是细胞生命活动所必需的,而交换方式与交换物质的类型密切相关。无机离子及小分子物质,如K+ 、Cl- 、Ca2+ 、Mg2+ 、水分子、单糖、氨基酸等可以自由地通过核膜。绝大多数大分子物质及一些小分子颗粒主要通过核孔复合体实现核-质之间的物质交换。核孔复合体通过主动运输和被动运输两种方式来实现核-质间的物质运输,具体转运方式取决于被转运物质的大小。离子、小分子及直径小于10nm 的物质原则上都可以被动运输的方式自由出入核孔。DNA 聚合酶、RNA 聚合酶、组蛋白等参与复制、转录、染色体的构建和核糖体前体组装等过程所需要的各种因子运输到核内,翻译所需要的RNA、组装好的核糖体亚单位等物质从细胞核运输到细胞质需要通过主动运输来实现。主动运输是核孔复合体最重要的功能之一,是一个信号识别与载体介导的过程,需要ATP,对温度敏感(图7G1G5)。
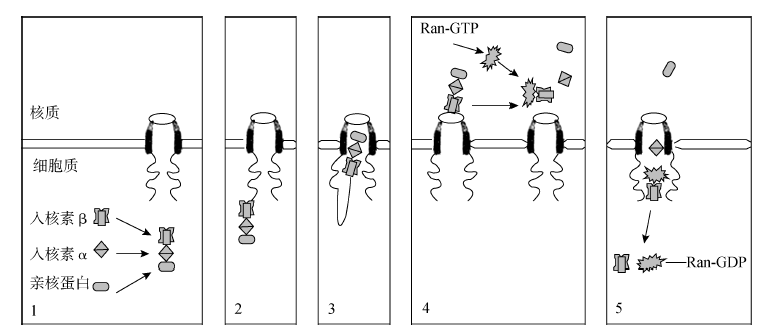
图7G1G5 亲核蛋白输入细胞核的过程
(三)参与细胞周期
核膜具有随着细胞周期的运转进行周期性解体和重建的特性。分裂间期核膜完整,分裂前期核膜崩解成单层膜泡,分裂末期核膜又将围绕染色体重新形成,如此周而复始。内核膜上存在LaminB受体,介导核膜与核纤层结合。在分裂前期结束时,核膜崩解为核膜小泡,核纤层被磷酸化解聚,LaminB与核膜小泡结合,LaminA和LaminC分散于细胞质中;在分裂末期,核膜小泡被引导至染色体表面,经膜融合构成围绕染色体的新核膜,核纤层蛋白去磷酸化重新组装;在细胞间期,核纤层为染色质提供锚定的位点,使染色质结合在核纤层内侧,阻碍染色质进一步螺旋化形成染色体。
(四)参与蛋白质的合成
外核膜的表面附着许多核糖体,可以进行蛋白质的合成。如抗体的形成首先出现在核膜外层。外核膜的附着核糖体也可以合成少量膜蛋白。

