
第一节 微 管
学习要求
知识内容
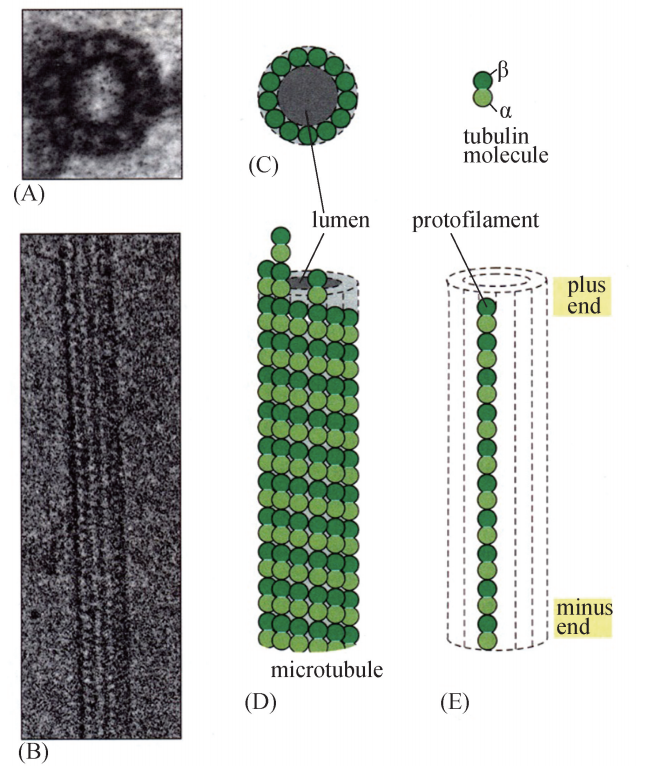
微管 (Microtubules)是外径25nm、内径15nm 的中空管 (图6G1G1),广泛分布 于真核生物各组织类型细胞中。例如,在神经组织内,主要的细胞类型是神经元,其 对于下游效应组织器官 (如肌肉组织)的调控是通过细胞胞体内产生的神经递质沿轴 突分泌至组织连接处、作用于靶点细胞而实现的,而轴突内用于长距离输送神经递质 的轨道系统主要由微管组装而成 (图6G1G2);位于气管或输卵管上皮细胞表面的纤毛结 构,也主要由微管组成,纤毛可以通过有规律的摆动,将气管内的黏液或输卵管内的 卵细胞分别朝着口腔或输卵管壶腹部方向推动。在红细胞及正在分裂的细胞内,微管 结构发挥维持细胞形状及协助将复制后的染色体平均分配至子细胞的作用。
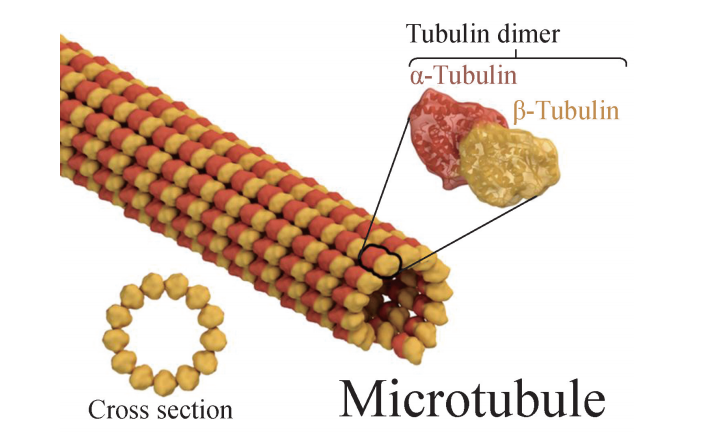
微管蛋白及其组成的微管原纤维和微管
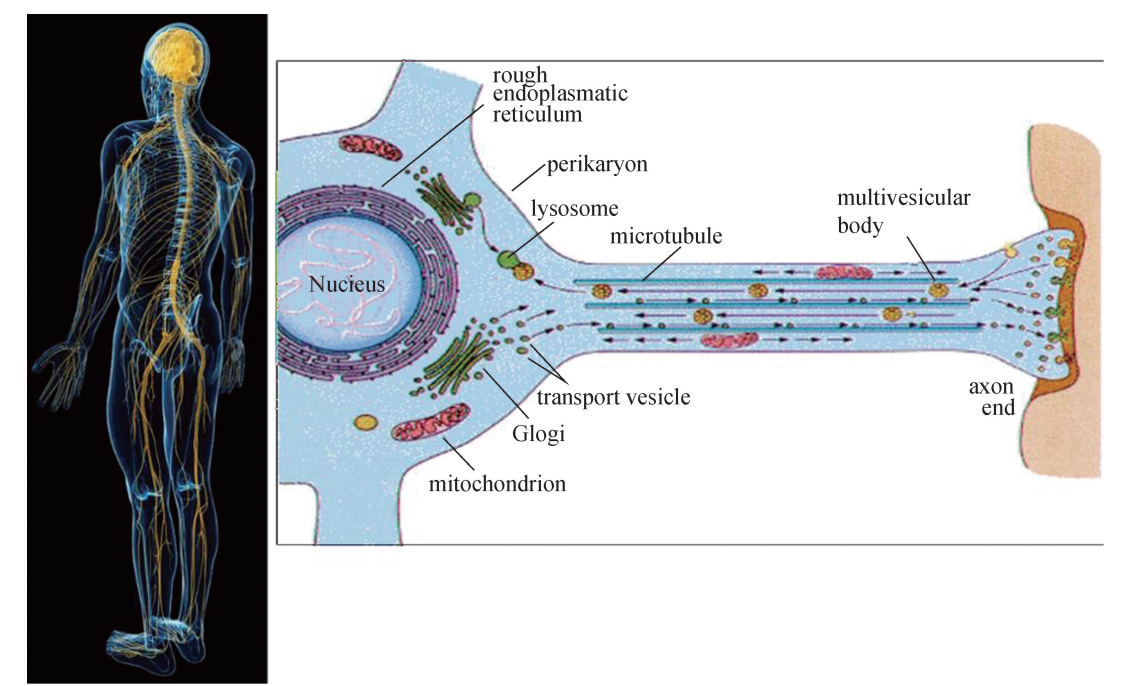
微管在组成神经系统的神经元轴突中的分布
一、 微管的结构、 化学组成与装配 (一) 微管的结构、 化学组成 细胞中 微 管 的 存 在 形 式 有 3 种:单 管 (singlet)、二 联 管 (doublet) 和 三 联 管 (triplet)(图6G1G3)。细胞中大部分微管都是单管,由13条原纤维 (protofilament)环 围而成,长短不一,主要由微管蛋白 (tubulin)和微管相关蛋白两类成分组成。二联 管主要构成纤毛和鞭毛的杆状部分,由 A 管和 B管组成,其中 A 管与单管结构相同, A 管有3条原纤维与B管共有;中心粒和鞭毛、纤毛的基体是三联管结构,由 A 管、B 管、C管组成,其中 A、B两管与二联管结构相同,C管有3条原纤维与 B管共有。
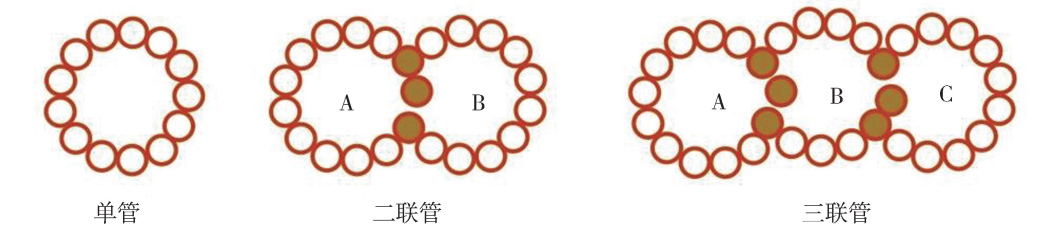
细胞中微管的3种存在形式
微管蛋白占总蛋白的80%,是一类酸性蛋白质。细胞质中的微管蛋白一般以异二 聚体 (heterodimer)的单体形式存在,构成异二聚体的单体蛋白质为 α- 微管 蛋 白 (α-tubulin)和β-微管蛋白 (β-tubulin),它们在空间结构上各有一个 GTP的结合 位点,相间排列成1条长链,称为原纤维,13根原纤维合拢形成一段微管。微管两端 都可延伸加长,但增长速度不同,速度快的一端为正端,另一端为负端,因而具有 极性。
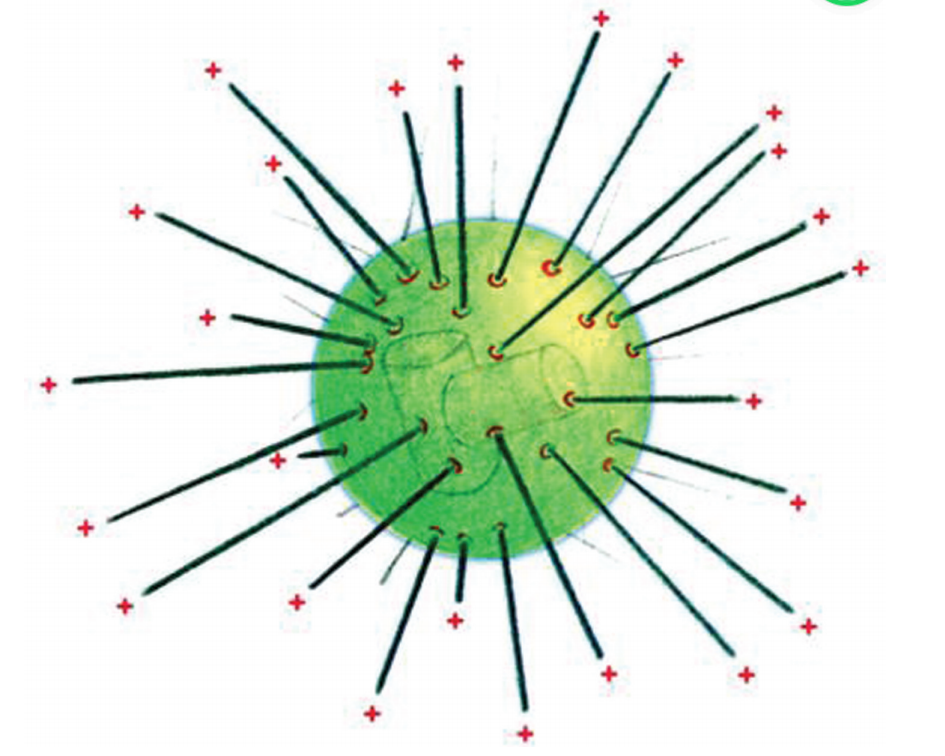
微管蛋白家族中的另一成员γ-微管蛋白 (图6G1G 4), 位 于 微 管 组 织 中 心 (microtubule organizing center,MTOC),分子量约为550000,多个 γ-微管 蛋白与其他蛋白质分子一起构成 γ-微管蛋白环状复 合物。尽管γ-微管蛋白只占微管蛋白总含量的不足 1%,但它参与纺锤体的形成,从而影响细胞分裂。 微管相关 蛋 白 (microtubuleassociatedproteins, MAPs)种类丰 富,是 微 管 结 构 和 功 能 必 需 的 成 分。 目前了解较多的 MAPs有以下四种:MAP1,MAP2, MAP4,Tau (作为一种可以同时连接微丝与微管的蛋 白质,也被列入 ABPs中的一种)。MAP1主要分布于 神经元的轴突与树突内,具有两个结构域,一个是碱 性的微管蛋白结合结构域,作为同带负电的微管蛋白
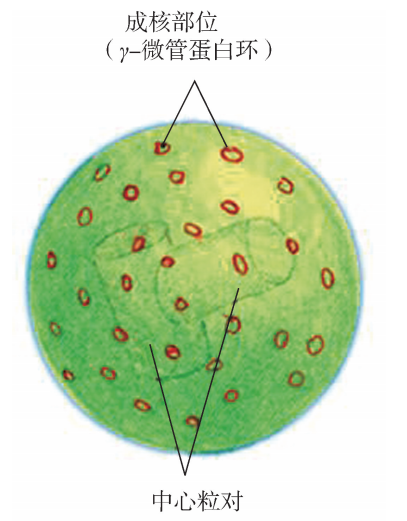
图6G1G4 γ-微管蛋白
结合的位点;另一个是酸性的外伸的结构域,起到控制微管在细胞内排列间距大小的 作用。MAP2,MAP4,Tau不同于 MAP1,它们均通过与微管结合的方式,达到稳定 微管结构和促进微管聚合的作用。 除了 MAPs这类内源性的、调控微管结构的分子外,自然界中也存在一些外源性 的、影响人体细胞微管聚合或解聚过程的物质分子。20世纪60至70年代,美国科学 家们在太平洋紫衫树皮中,发现并提纯了一种能够抑制肿瘤细胞生长,名为紫杉醇 (PaclitaxelorTaxol)的成分,其抗癌作用的分子基础在于通过结合在微管管腔的内表 面,使微管难以解聚,使正在进行有丝分裂的细胞中的纺锤体结构不能够将复制后的 遗传物质平均分配到两个子细胞中,最终使活跃癌细胞的增殖能力大幅下降,达到抑 制肿瘤生长的目的。临床上,紫杉醇主要用于治疗乳腺癌和肺癌。另外一种用于治疗 肺癌的化疗药长春新碱 (Vinblastine),也是通过作用在细胞骨架的微管结构来达到抑 制肿瘤细胞生长的目的的。但是,与紫杉醇稳定微管结构的作用相反,该化疗药可以 与微管的单体结合,从而阻止其聚合成原纤维乃至微管结构。同样可以通过阻止游离 微管蛋白异源二聚体组装成微管结构的一种临床抗炎药物秋水仙素 (Cochicine),则是 治疗急性痛风发作的常用药。 (二) 微管的装配 根据细胞的生理需要,微管蛋白通过聚合或解聚,引起微管装配。根据微管动态 存在的时间长短,细胞中有两类微管,一种微管在细胞中存在时间很短,发生快速组 装和去组装,称为动态微管 (dynamicmicrotubule),如纺锤体;另一些微管存在时间 相对较长,称为稳定微管 (stablemicrotubule),构成一些特化的细胞结构,如纤毛。 作为对周围环境变化的反应,微管的稳定性可以改变。 1 微管的体外装配 在适当条件下,微管能在体外自我装配。微管的装配是一个 复杂而有序的过程,微管蛋白浓度、pH 值和温度都会影响装配速度。微管的装配分为 成核期、聚合期和稳定期3个时期。 (1)成核期 (nucleationphase):首先,α-微管蛋白和β-微管蛋白形成长度为 8nm 的异二聚体。异二聚体先沿纵向聚合形成1个短的寡聚体 (oligomer)核心,这个 寡聚体可能是不够稳定的。然后再以这个寡聚体核心为基础,经过侧面增加二聚体而 扩展为弯曲的片状 (sheet)结构,这种片状结构的稳定性大大提高。当片状带加宽至 13根原纤维时,即合拢形成一段微管 (图6G1G5)。此时由于是微管聚合的开始,速度 缓慢,因此又称为延迟期。 (2)聚合期 (polymerizationphase):也称作延长期,二聚体以较快的速度从两端 加到已经形成的微管上,此时微管不断地延长。 (3)稳定期 (steadystatephase):随着细胞质中的游离微管蛋白的浓度下降,当 达到临界浓度时,微管聚合与解聚的速度达到平衡,此时微管组装与去组装的速度相 等,微管的长度相对恒定。
微管的成核期
2 微管的体内装配 活细胞内的微管组织中心在微管体内装配过程中起着重要作 用,包括中心体、基体和着丝点等,它们提供了微管组装所需要的核心。位于微管组 织中心的γ-微管蛋白环复合物作为微管成核中心与微管的负端结合,指导微管蛋白二 聚体结合上去,并按照体外装配的规律进行微管的组装。先组装出 (-)极,然后开 始生长,因此中心体周围的微管 (-)极指向中心体,(+)级远离中心体 (图6G1G6)。
微管的体内装配
二、 微管的功能 1 维持细胞形态 微管和其他细胞骨架成分协同维持细胞形态。微管具有一定的 强度,能够抗压和抗弯曲,给细胞提供机械支持力,是支撑和维持细胞形状的主要物 质。典型的如神经细胞长长的突起就是靠成束的微管支撑。微管对细胞内细胞器的位 置固定也有重要作用。 2 参与物质运输 细胞内的细胞器移动和胞质中的物质转运都和微管有着密切的 关系,具体功能由马达蛋白来完成。马达蛋白是指介导细胞内物质沿细胞骨架运输的 蛋白,主要分三大类:动力蛋白 (cytoplasmicdynein)、驱动蛋白 (kinesin)、肌球蛋 白 (myosin)。前两类负责将物质沿微管运输 (图6G1G7),后一类负责将物质沿微丝运 输。典型的如变色龙体表内含特化的色素细胞,在神经控制下,色素细胞中的色素颗 粒可在几秒内沿微管运动,使皮肤变色 (图6G1G8)。
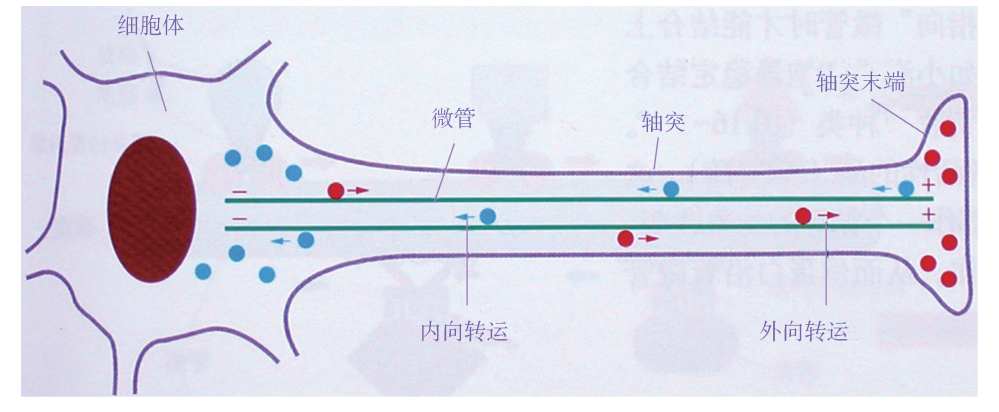
图6G1G7 神经轴突中沿微管的转运 (红色:驱动蛋白;蓝色:动力蛋白)
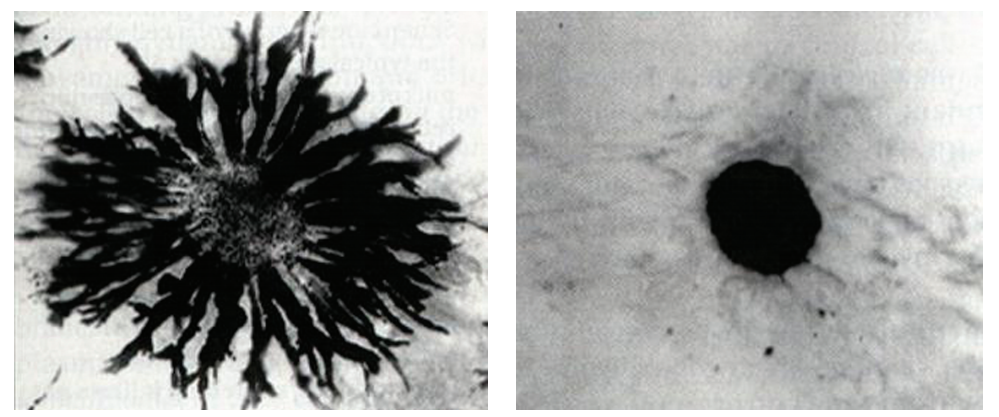
色素颗粒沿微管移动
3 参与细胞分裂 微管组成细胞的中心粒,中心粒是短管状小体,直径 160~ 260nm,长160~560nm。细胞中的中心粒成对存在且相互垂直,它们连同其周围物质 构成光镜下所见的中心体 (centrosome) (图6G1G9)。横切面可见其圆柱状小体的壁由 9组三 联 管 斜 向 排 列 呈 风 车 状 包 围 而 成,为 “9+0” 的 结 构。中 心 粒 在 细 胞 中 起 MTOC的作用,在细胞间期,中心体组织形成胞质微管,在细胞分裂期,组织形成纺 锤体,参与细胞有丝分裂。
图6G1G9 中心体模式
4 参与细胞运动 纤毛和鞭毛是真核细胞表面伸出的与运动有关的特化结构,通 常将少而长的称为鞭毛,短而多的称为纤毛。极少数细胞通过鞭毛和纤毛进行运动, 如精子依靠鞭毛的摆动进行游动,呼吸道上皮细胞依靠纤毛的摆动向气管外排送痰液。 纤毛 和 鞭 毛 的 主 要 成 分 是 微 管, 两 者 在 结 构 上 同 源, 都 是 由 细 胞 膜 包 绕 轴 丝 (axoneme)构成的。轴丝为 “9+2”结构,即9组二联管环绕1对中央单管 (图6G1G 10)。中央单管由中央鞘包围,单管之间有横桥相连。相邻的二联管之间也有蛋白质相 连,并且八管伸出两条动力蛋白臂,指向邻近二联管的 B 管。动力蛋白臂能引起二联 管之间的相互滑动,导致纤毛和鞭毛摆动。中央单管呈放射状发出放射幅条 (radial spokes),伸向周围的 A 管。轴丝是以纤毛和鞭毛的基体作为 MTOC 组装而成的。基 体的结构与中心粒一样,呈 “9+0”排列,即含9组三联管,无中央微管。在基体和 轴丝之间有一段转换区。

纤毛的结构

