
第二节 细胞呼吸与能量转换
学习要求
知识内容
一、 细胞呼吸
较高等的动物都能依靠呼吸系统从外界吸取 O2 并排出 CO2,从某种意义上说细胞 中也存在有这样的呼吸作用,即在细胞内特定的细胞器 (主要是线粒体)内,在 O2 的 参与下,分解各种大分子物质产生 CO2;与此同时,分解代谢所释放出的能量储存于 ATP中,这一过程称为细胞呼吸 (cellularrespiration),也称为生物氧化 (biological oxidation)或细胞氧化 (cellularoxidation)。细胞呼吸是细胞内提供生物能源的主要途 径,它的化学本质与燃烧反应相同,最终产物都是 CO2 和 H2O,释放的能量也完全相 等。但是,二者所进行的方式却大不相同。细胞呼吸是在细胞内进行的,反应条件温 和 (在体温及近于中性pH 值条件下进行)。细胞呼吸所包含的化学反应几乎都是在线 粒体中进行的一系列由酶系所催化的氧化还原反应,能量逐步放出,且放出的能量一 般是以化学能的方式储存在 ATP中,这样所产生的能量利用率高,不会像物质在体外氧化那样,能量以光和热的形式瞬间放出。 ATP是一种高能磷酸化合物,细胞呼吸时,释放的能量可通过 ADP 的磷酸化而 及时储存于 ATP的高能磷酸键中作为备用;反之,当细胞进行各种活动需要能量时, 又可去磷酸化,断裂一个高能磷酸键以释放能量来满足机体需要。ATP的放能、储能 反应简式如下:
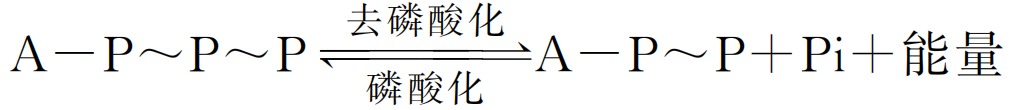
随着细胞内不断进行的能量释放和储存,ATP 与 ADP 不停地进行着互变。因为 ATP是细胞内能量转换的中间携带者,所以被形象地称为 “能量货币”。ATP是细胞 生命活动的直接供能者,也是细胞内能量获得、转换、储存和利用等环节的联系纽带。 ATP中所携带的能量来源于糖、氨基酸和脂肪酸等的氧化,这些物质的氧化是能 量转换的前提。以葡萄糖氧化为例,细胞有氧呼吸产生 ATP的过程大体分为四个主要 步骤:①糖酵解;②三羧酸循环;③电子传递;④ATP合成 (图5G2G1)。除糖酵解在 细胞质中完成外,其他3个阶段均在线粒体内完成。
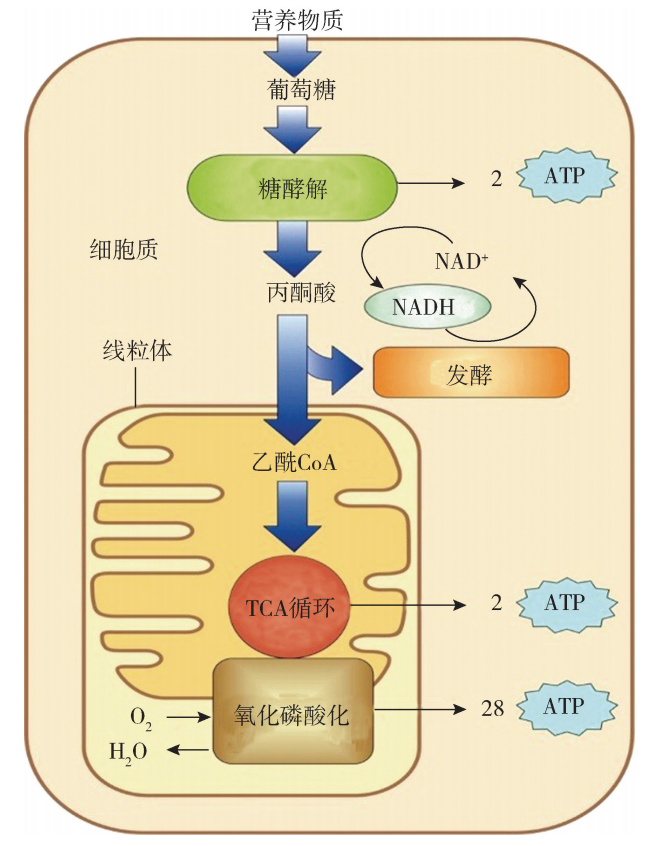
细胞呼吸的四个主要步骤
二、 细胞质中的糖酵解 从葡萄糖分解为丙酮酸的过程中,不需要氧的参与且在细胞质中进行,故称为糖酵解 (glycolysisi)。经过糖酵解过程的十几步酶促化学反应,1分子葡萄糖分解生成2 分子丙酮酸,2分子 NADH+H+ 和2分子 ATP。反应式如下: C6H12O6+2NAD+ +2ADP+2Pi→2CH3COCOOH+2NADH+2H+ +2ATP ATP是通过底物水平磷酸化产生的。底物水平磷酸化 (substrateGlevelphosphoG rylation)是指高能底物水解放能,直接将高能磷酸基从底物转移到 ADP上,使 ADP 磷酸化生成 ATP(图5G2G2)。在没有线粒体、不能进行有氧氧化的细胞 (如红细胞), 糖酵解是一条重要的产能途径。它提供的能量虽然少,但产能速度快,能满足能量的 应急需求。剧烈运动时肌肉细胞的能量供应就依靠糖酵解,剧烈运动引起的肌肉酸痛, 是由于缺氧状态下糖酵解产生的丙酮酸还原为乳酸,堆积在肌组织中所致。
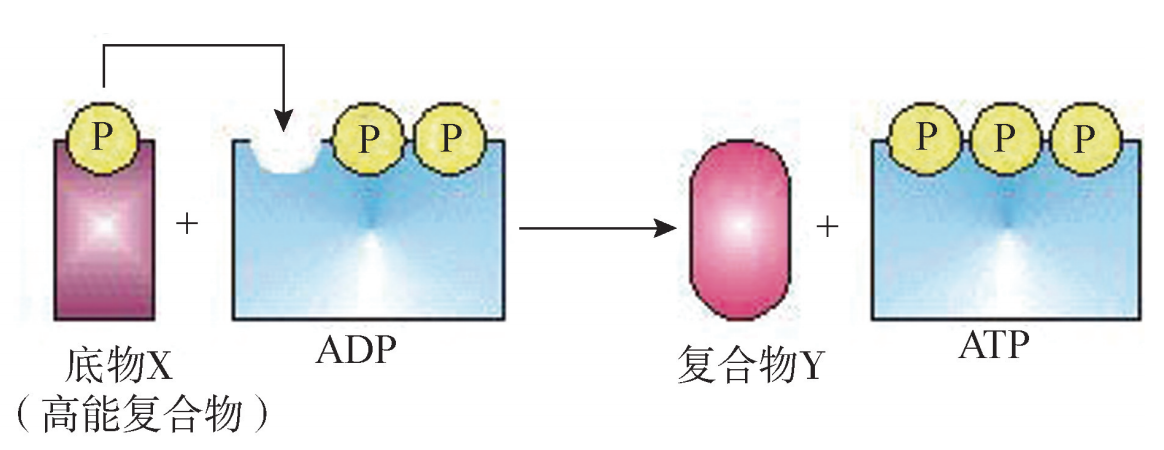
底物水平磷酸化过程
在供氧充足情况下,糖酵解产物丙酮酸和 “NADH+H+ ”从细胞质基质中进入线 粒体,在线粒体基质腔内丙酮酸脱氢酶系作用下,丙酮酸与辅酶 A(HSCoA)反应, 生成乙酰辅酶 A;同时,烟酰胺腺嘌呤二核甘酸 (NAD+ ) 作为受氢体被还原为 NADH+H+ ,脱下的羧基形成 CO2。此过程称为乙酰辅酶 A(CH2COSCoA)形成。 反应式如下:
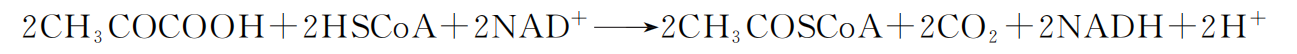
三、 线粒体中 ATP合成的三部曲 (一) 三羧酸循环 在线粒体基质腔中,乙酰辅酶 A 与草酰乙酸结合成柠檬酸进入柠檬酸循环。由于 柠檬酸有三个羧基,故柠檬酸循环也称为三羧酸循环 (tricarboxylicacidcycle,TCA cycle,图5G2G3)。三羧酸循环包括一系列酶促的氧化脱氢和脱羧反应,循环的末端又 重新生成草酰乙酸。每一次循环,消耗3个 H2O,生成4对 H 和2分子 CO2,底物水 平磷酸化生成1分子 GTP(可变为1分子 ATP),脱下的4对 H 有3对以 NAD+ 为受 氢体,形成3分子 NADH+H+ ,受氢体 FAD(黄素腺嘌呤二核苷酸)接受另外1对 H 后转变为还原态 FADH2。总反应式如下:
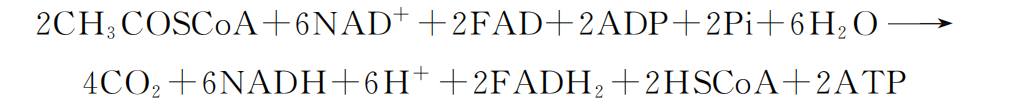
至此,1分子葡萄糖经过分解共形成了10分子 NADH+H+ 、2分子 FADH2、4 分子 ATP和6分子 CO2。
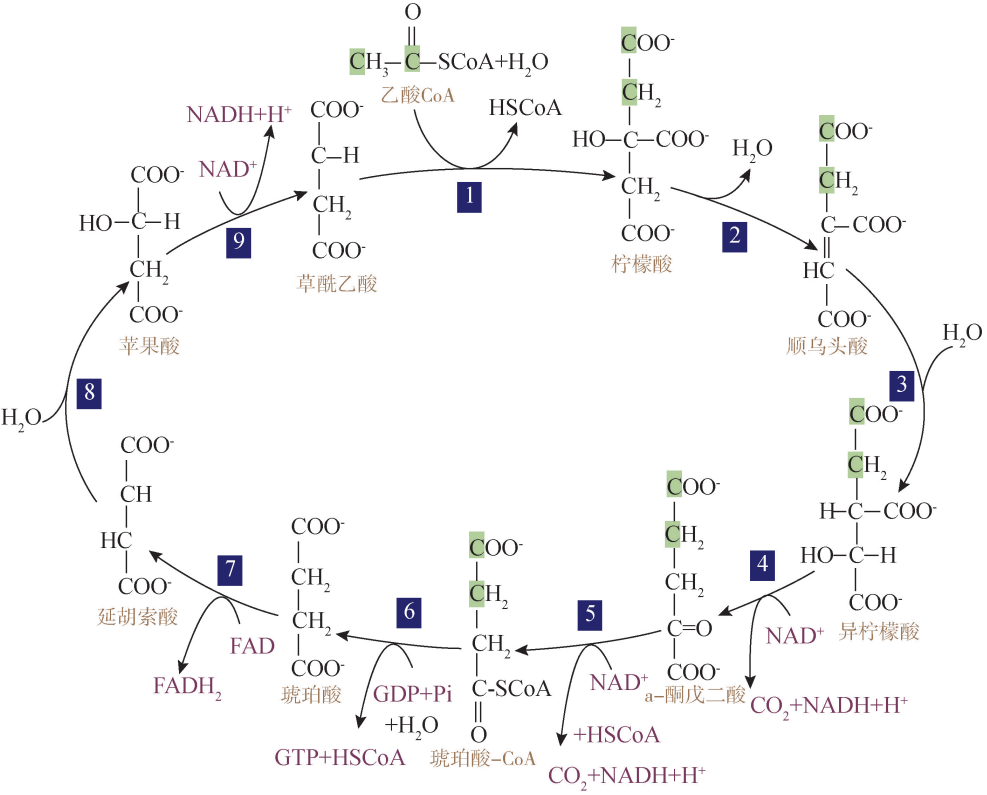
三羧酸循环
三羧酸循环是各种有机物进行最后氧化的过程,也是各类有机物相互转化的枢纽。 除了丙酮酸外,一些氨基酸也从细胞质进入线粒体,并进一步转化成乙酰 CoA 或三羧 酸循环的其他中间体。三羧酸循环的中间产物可用来合成包括氨基酸、卟啉及嘧啶核 苷酸在内的许多物质。只有经过三羧酸循环,有机物才能进行完全氧化,提供远比糖 无氧酵解所能提供的多得多的能量,供生命活动的需要。
(二) 呼吸链
电子传递 (electrontransport)化磷酸化 (oxidativephosphorylation)是能量 转换的主要环节。
高能电子沿线粒体内膜的电子传递链 (electrontransportchain)或 呼吸链 (respiratorychain)传递过程中释放能量,释放的能量在基粒内通过氧化磷酸 化生成 ATP。因此,电子传递链是电子传递的结构基础,基粒是 ATP合成的场所 (图 5G2G4)
经过上述3个环节,供能物质的大部分能量已经以高能电子形式转移至 NADH+ H+ 、FADH2,经过电子传递链的电子传递和基粒的氧化磷酸化作用,NADH、 FADH2 拥有的能量逐步释放并转移至 ATP中。 具有递氢、递电子作用的一系列氢载体和电子载体 (electroncarrier),在线粒体 内膜上有序排列,构成相互关联的链状,称为电子传递链。该体系最终以氧作为电子

线粒体内膜电子传递复合物的排列及质子与电子传递示意图
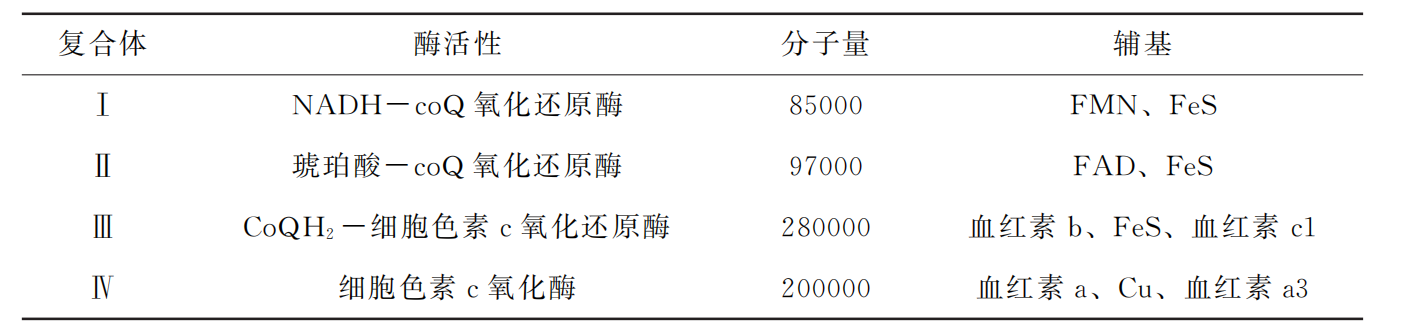
接受体,与细胞摄氧有关,故又称为呼吸链。除泛醌 (ubiquinone,UQ)和细胞色素c (Cytc)外,电子传递链的其他成员均是内膜的整合蛋白质。UQ 是脂溶性醌类化合物, 可在脂双层中从膜的一侧向另一侧移动;Cytc是膜周边蛋白质,可在膜表面移动。电 子传递链的组分在线粒体内膜上分别组成Ⅰ、Ⅱ、Ⅲ、Ⅳ四个酶复合体,它们是线粒 体内膜的整合蛋白 (表5G2G1)
线粒体呼吸链的组分
(三) ATP合成 1 基粒催化 ATP的合成 线粒体内膜 (包括嵴)的内表面附有许多圆球形基粒。 基粒由头部、柄部和基底部 3 部分组成:头部呈球形,直径 8~9nm;柄部直径约 4nm,长4头部与柄部相连凸出在内膜表面,柄部则与嵌入内膜的基片相 连。进一步研究表明,基粒是将呼吸链电子传递过程中所释放的能量 (质子浓度梯度 和电位差)用于使 ADP 磷酸化生成 ATP 的关键装置,是由多种多肽构成的复合体, 其化学本质是 ATP 合酶 (ATPsynthetase,ATPase)或 ATP 合酶复合体,也称为 F0F1ATP合酶 (图5G2G5)
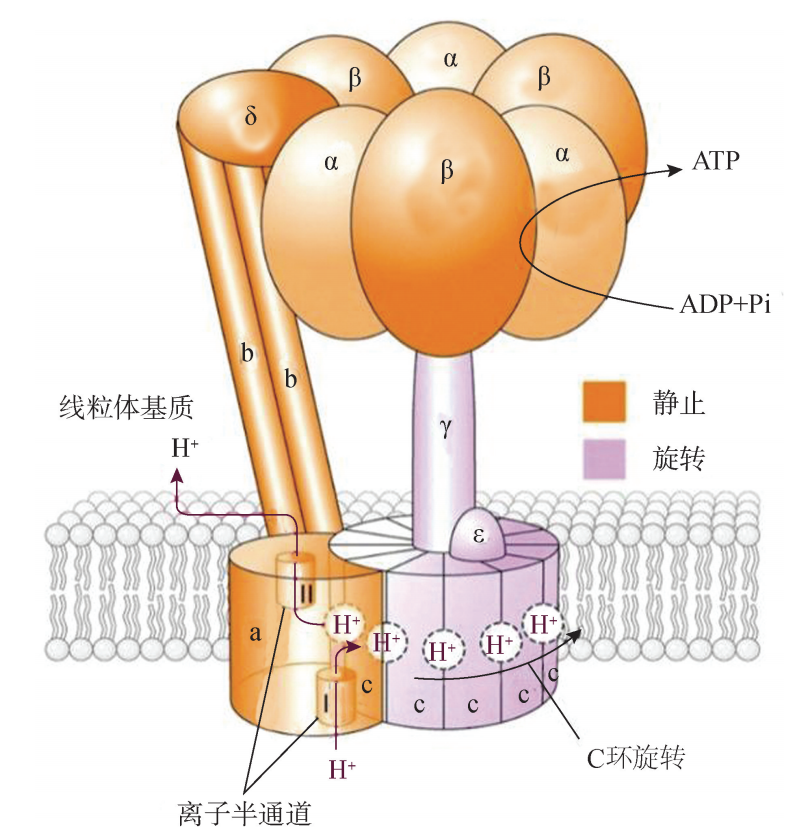
线粒体的 ATP合成酶的基本结构
(显示其头部、柄部和基底部3部分组成)
线粒体 ATP合成酶是生物能量转换的核心酶,参与氧化磷酸化,在跨膜质子动力 势能推动下催化合成 ATP。1997年诺贝尔化学奖授予了三位从事 ATP合成酶研究的 科学家,说明 ATP合成酶研究工作的意义。不同来源的 ATP合成酶基本有相同的亚 基组成和结构,都是由许多亚基装配形成的。 (1)头部:一般认为是 ATP复合物 (也称为偶联因子 F1),它位于线粒体内膜的 基质侧表面,由5种亚基组成,是9聚体 (α3β3γδε),含有 ATP合成酶活性,其中α 和β亚基上有 ADP和 ATP的结合位点;β亚基有催化活性,称为催化亚基;γ亚基可 调节质子从 F0 蛋白向 F1 蛋白的流动,起着 “阀门”的作用。 (2)柄部:位于头部 (F1)和基底部 (F0)之间,也起着调节质子流的作用,柄 部由三种蛋白组成,其中一种对寡霉素敏感,称为寡霉素敏感蛋白 (oligomycinsensiG tivityconferringprotein,OSCP),因此也将柄部和底部合称为 F0,ATP 合成酶复合 物主要指 F0-F1 蛋白。 (3)基底部:即 F0,为疏水的内在蛋白质,镶嵌在线粒体内膜中,呼吸链围绕其 周围,它由4种亚基组成,这些亚基在内膜中形成了跨膜的质子通道,质子从内膜外 侧径柄部流向 F1 蛋白。F0 中的0表示对寡霉素敏感的部位。
2 氧化过程伴随着磷酸化的偶联
经糖酵解和三羧酸循环产生的 NADH 和 FADH2 是两种还原性的电子载体,它们所携带的电子经线粒体内膜上的呼吸链逐级定 向传递给 O2,本身则被氧化 (图5G2G6)。由于电子传递所产生质子 (H+ )浓度梯度和电位差,其中所蕴藏的能量被 F0F1ATP合酶用来催化 ADP磷酸化而合成 ATP。氧化 和磷酸化相耦联,称为氧化磷酸化。因此,线粒体内膜上的电子传递链可认为是 “放 能装置”,而基粒则是 “换能装置”,“放能”与 “换能”相耦联。
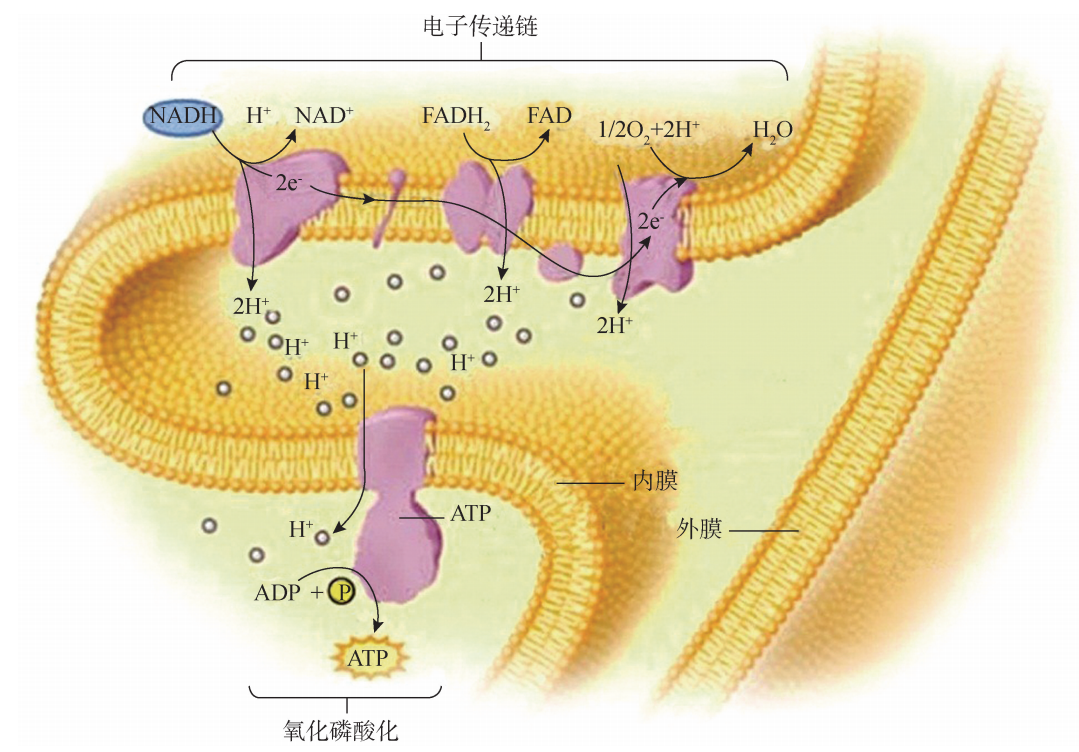
电子传递与氧化磷酸化过程
氢载体 NADH 和 FADH2 进入电子传递的部位不同,所释放的能量也有差异。1 分子的 NADH 经过电子传递可形成2分子 ATP,而1分子 FADH2 经过电子传递可形成1分子 ATP。电子传递链的其他部位释放的能量不足以形成 ATP,只能以热量 形成耗散。因此,物质氧化分解经电子传递所释放的能量并不是全部形成 ATP。 综上所述,1分子葡萄糖完全分解形成的10分子 NADH 和2分子 FADH2,经氧 化磷酸化生成28个 ATP,糖酵解过程底物水平磷酸化产生2个 ATP(细胞质中),三 羧酸循环过程底物水平磷酸化产生2个 ATP。所以,1分子葡萄糖完全氧化产生32个 ATP。12个 H 解离形成12H+ 与电子传递链终端产生的 O2- 结合生成水,即12H+ + 6O2- →6H2O。 氧化磷酸化所需要的 ADP和Pi由细胞质基质输入到线粒体基质,而线粒体内合成 的 ATP则输送到细胞质基质,供细胞的生命活动利用。可是线粒体内膜具有高度不透 性,因此,这些物质进出线粒体需要依靠专门的结构。线粒体内膜上有一些专一性转 运蛋白与这些物质进出线粒体有关,例如,其中的一种为腺苷酸转移酶能利用内膜内 外 H+ 梯度差把 ADP和 Pi运进线粒体基质,而把 ATP输往线粒体外。
3 电子传递时 H+ 穿膜形成电化学梯度 关于电子传递同磷酸化的耦联机制至今尚未彻底阐明,曾先后有过许多假说,目前被广泛接受的是英国化学家 P (1961)提出的化学渗透假说 (chemiosmoticcouplinghypothesis)。该假说认为氧化磷 酸化耦联的基本原理是电子传递中的自由能差造成 H+ 穿膜传递,暂时转变为横跨线粒 体内膜的电化学质子梯度 (electrochemicalprotongradient)。然后,质子顺梯度回流 并释放出能量,驱动结合在内膜上的 ATP合酶,催化 ADP磷酸化合成 ATP。这一过 程可综合如下:①NADH 或 FADH2 提供一对电子,经电子传递链,最后为 O2 所接 受;②电子传递链同时起 H+ 泵的作用,在传递电子的过程中伴随着 H+ 从线粒体基质 到膜间腔的转移;③线粒体内膜对 H+ 和 OH- 具有不可透性,所以随着电子传递过程 的进行,H+ 在膜间腔中积累,造成了内膜两侧的质子浓度差,从而保持了一定的势能 差;④膜间腔中的 H+ 有顺浓度返回基质的倾向,能借助势能通过 ATP酶复合体F0 上 的质子通道渗透到线粒体基质中,所释放的自由能驱动 F0F1ATP合酶合成 ATP。 化学渗透假说有两个特点,一是需要定向的化学反应;二是突出了膜的结构。该 学说可以解释氧化磷酸化过程中的许多特性,也得到了很多实验结果的支持。但是也 仍存在一些难以用化学渗透假说解释的实验结果,因此还必须不断地修改和完善。相 继有人提出了一些新的理论,包括变构假说、碰撞假说等,但都存在一定的问题。

