
第五节 囊泡与囊泡转运
学习要求
知识内容
囊泡 (vesicle)也称为小泡,由一层单位膜包裹而成的临时性动态结构,是真核细 胞中常见的膜泡结构,是内膜系统中不可或缺的结构功能组分。囊泡运输 (vesicular trafficking)是指将大分子物质和颗粒物质在细胞内转运过程中完成膜本身的融合、重 组和移位的过程。囊泡的形成和出现,都会伴随着细胞内物质的定向运输过程。
一、 囊泡的种类与来源
囊泡是内膜系统中重要的整体功能结构组分之一,其并不是一种相对稳定的细胞 内固有结构,而是细胞内物质定向运输的载体和功能表现形式。内膜系统各个部分之间的物质传递常常通过膜泡运输方式进行,如从内质网到高尔基体,高尔基体到溶酶 体,细胞分泌物的外排,都要通过过渡性小泡进行转运。根据大量研究推测,承担细 胞内物质定向运输的囊泡类型至少有10种以上,但是目前了解较多的有网格蛋白有被 囊泡 (clathrinGcoatedvesicle)、COPⅠ有被囊泡 (COPⅠcoatedvesicle)和 COPⅡ有 被囊泡 (COPⅡcoatedvesicle)三种类型。
(一) 网格蛋白有被囊泡 网格蛋白有被囊泡产生于反面高尔基复合体的网络结构,也可由受体介导的细胞 内吞作用形成。由高尔基复合体产生的网格蛋白小泡,介导蛋白质从反面高尔基体网 状结构向胞内体、溶酶体或细胞膜运输。在受体介导的胞吞作用过程中,介导物质从 细胞膜向细胞质或从胞内体运输到溶酶体。 网格蛋白有被囊泡的直径一般在50~100nm,主要有以下两个特征:第一,网格 蛋白 (clathrin)纤维构成外被的框架结构,也因此而得名;第二,在网格蛋白结构外 被与囊膜之间的20nm 间隙中充填有衔接蛋白 (adaptin),介导网格蛋白与囊膜跨膜蛋 白受体的连接,提高了囊泡的表面张力。在网格蛋白的形成过程中,发动蛋白 (dyG namin)也发挥着重要作用。发动蛋白在细胞质中结合并水解 GTP,当囊膜形成时, 在芽生囊膜以环状聚合在颈部。随着其对 GTP的水解,环状的发动蛋白向心收缩,囊 泡断裂分离,脱去网格蛋白外被,转化为无被小泡,开始向靶膜转运 (图4G5G1)。
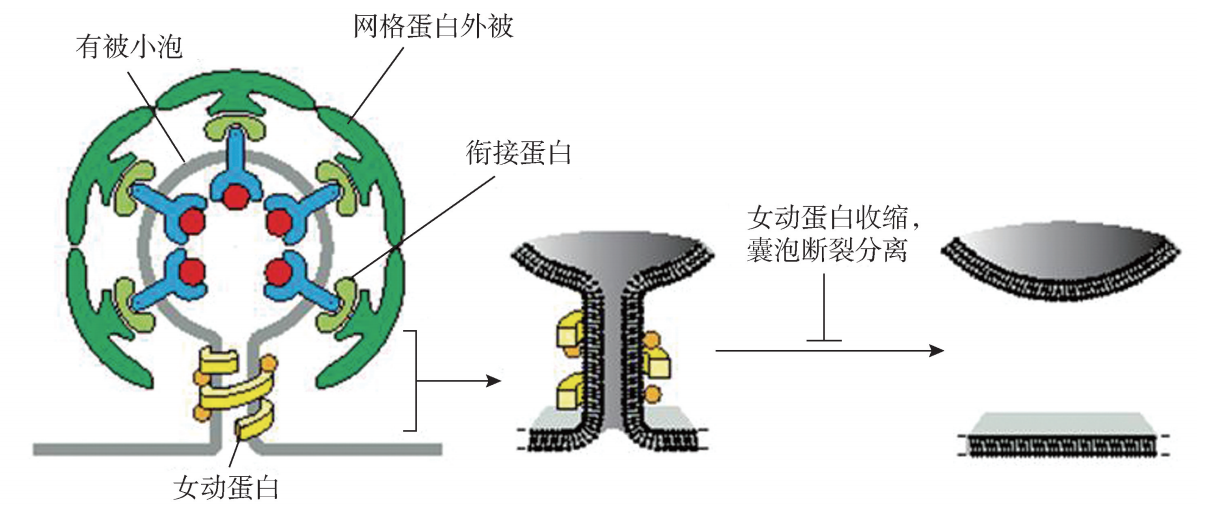
图4G5G1 网格蛋白有被囊泡结构示意
(二) COPⅠ有被囊泡
COPⅠ有被囊泡产生于高尔基复合体顺面,囊泡表面有衣被蛋白Ⅰ覆盖,主要负 责内质网逃逸蛋白 (escapedprotein)的捕捉、回收转运以及高尔基体膜内蛋白的逆向 运输。有证据表明,COPⅠ有被囊泡也能够行使从内质网到高尔基体的顺向转移。顺 向转移一般不能直接完成,通常需要内质网-高尔基体中间体这一中间环节的中转。 COPⅠ有被囊泡的外被蛋白是由7个亚基组成 (α、β、γ、δ、ε、ζ等)的多聚体, 其中α蛋白 (也称为 ARF蛋白)类似于 COPⅡ中的 Sar蛋白亚基,是一种 GTP结合蛋白,可调控外被蛋白复合物的聚合、装配及膜泡转运。 COPⅠ囊泡的形成过程包括:①GTP与 ARF 复合体的形成,即游离于细胞质中 的非活化状态的 ARF蛋白与 GDP解离并与 GTP结合;②GTP-ARF 复合体与高尔 基体膜上的 ARF受体识别、结合;③COPⅠ蛋白亚基聚合,与 ARF一起和高尔基体 囊膜表面其他相关蛋白结合,诱导转运囊泡生芽。当 COPⅠ有被囊泡从高尔基体顺面 囊膜断离下来,COPⅠ蛋白随即解离,COPⅠ有被囊泡转化为无被小泡运向靶膜。
(三) COPⅡ有被囊泡
COPⅡ有被囊泡由粗面内质网产生,因囊泡表面有衣被蛋白Ⅱ覆盖而得名,主要 介导内质 网 到 高 尔 基 体 的 物 质 转 运。研 究 发 现,COPⅡ 外 被 蛋 白 由 5 种 (SarⅠ、 sec23/sec24、sec13/sec31、sec6和sec12)亚基组成,其中 SarⅠ属于 GTP结合蛋白, 它可通过与 GTP或 GDP 的结合,调节囊泡外被的装配与去装配。SarⅠ蛋白亚基与 GDP结合,使之处于非活性状态;而与 GTP结合时,则激活SarⅠ蛋白导致其与内质 网膜结合,同时引发其他蛋白亚基在内质网膜上的聚合、装配、出芽及断离,形成 COPⅡ有被囊泡。试验证明,当 COPⅡ有被囊泡在内质网生成之后,在向高尔基体的 转移途中,常常多个囊泡彼此先行融合,形成内质网-高尔基体中间体,然后沿着微 管运行,到达高尔基体的顺面。COPⅡ有被囊泡在抵达靶标之后,与靶膜融合之前, 即由结合的 GTP水解,产生SarGGDP复合物,促使囊泡外被蛋白发生水解去装配,囊 泡脱被成为无被运输小泡。
二、 囊泡转运
囊泡转运是指囊泡以出芽的方式,从一种细胞器膜形成后按一定路径到达靶细胞 器,通过对靶细胞器进行识别而卸载其内容物的过程。因此,囊泡转运实现了真核细 胞中内膜系统各个部分之间的物质运输,同时也担负细胞与细胞外环境之间的物质和 信息交流。不同类型和来源的囊泡介导不同物质的运输,它们必须沿着正确的路径, 以特定的方式运输,具体有以下特点:
1 囊泡转运是细胞物质定向运输的基本途径
囊膜均来自细胞器膜,囊泡是由细 胞器膜以外突或内陷的形式芽生而成。囊泡的形成和产生过程,伴随着物质的转运; 囊泡的运行轨迹及归宿,取决于其所转运物质的定位去向。例如,细胞通过胞吞作用 摄入的外源性物质,总是以网格蛋白有被小泡的形式,自外向内从细胞膜输送至细胞 内体或溶酶体;而在细胞内合成的各种外输性蛋白及颗粒物质,首先是先进入内质网, 然后经过一系列的修饰、加工和质量检查后,以 COPⅡ有被囊泡的形式输送到高尔基 体,再直接地或经由溶酶体到达细胞膜,最终以胞吐作用分泌释放至细胞外。一旦内 质网驻留蛋白或折叠错误的外输性蛋白从内质网逃逸外流,在它们进入高尔基体后就 会被捕捉、回收,并由 COPⅠ有被囊泡遣送返回到内质网中。因此,囊泡转运介导物 质的双向运输,它不仅是细胞内外物质交换和信号传递的重要途径,也是细胞物质定 向运输的基本形式。
2 特异性识别融合是囊泡准确转运的基本保障
囊泡与靶膜的相互识别是它们之 间进行融合的前提,这种识别机制与囊泡表面的特异性标记分子和靶膜上的相应受体 密切相关。目 前 普 遍 认 为,所 有 转 运 囊 泡 及 细 胞 器 膜 表 面 都 各 自 带 有 一 套 特 有 的 SNAREs (可溶性 N-乙酰马来酰亚胺敏感因子结合蛋白受体)互补序列,位于运输小 泡上的叫v-SNAREs,位于靶膜上的叫t-SNAREs,两者互为识别,特异互补。两 种SNAREs都具有一个螺旋结构域,能相互缠绕形成跨SNAREs复合体,形成锁-钥 嵌合式的结构,并通过这个结构将运输小泡的膜与靶膜拉在一起,实现小泡特异性停 泊和融合,保证囊泡物质定向运输和准确卸载。目前,已经分离、鉴定的细胞内定位 的这两种蛋白家族成员多达10余种。
3 囊泡转运是驱动细胞膜及内膜系统功能结构转换和代谢更新的桥梁
囊泡转运 伴随着物质运输和膜的流动,当内质网出芽产生的转运囊泡汇至高尔基体时,其囊膜成 为高尔基体顺面囊膜的一部分;当高尔基体反面囊膜出芽产生和分化出不同的分泌囊泡 时,可直接运送至细胞膜,或经溶酶体最终流向和融入细胞膜;细胞膜通过内陷形成的网格 蛋白有被囊泡,则以胞内体或吞噬体的形式与溶酶体发生融合转换。由此可见,囊泡转运承 载和介导细胞物质定向运输的同时,又不断地被融合、更替和转换,从一种细胞器膜到另一 种细胞器膜,形成了一个高度有序、源源不断的膜流 (图4G5G2)。
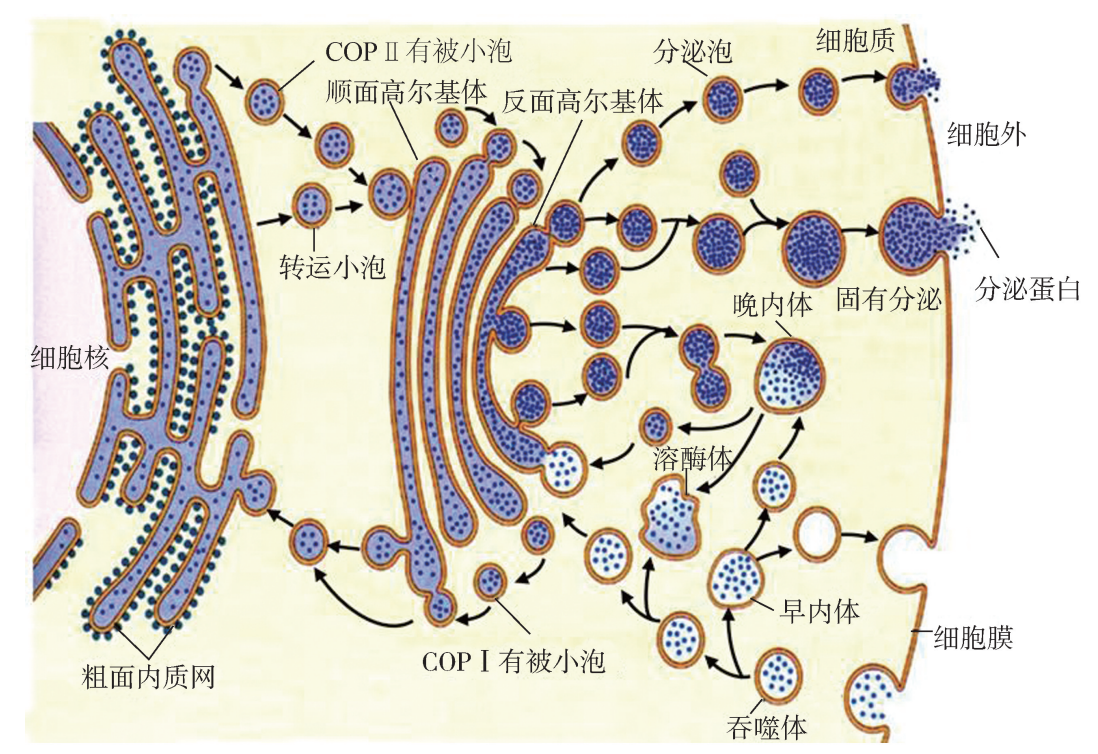
囊泡转运过程示意

