
学习要求
知识内容
内膜系统 (endomembrane system)是指位于细胞质内,在结构、功能乃至发生上相互密切关联的膜性结构细胞器的总称。主要包括:内质网、高尔复合基体、溶酶体、 过氧化物酶体、各种转运小泡以及核膜 (图1)。
内膜系统的功能意义:
1. 内膜系统是真核细胞与原 核细胞之间在形态、结构上相互区别的重要标志。
2.细胞内部结构不断分化完善,各种生理功能逐渐提高
3.内膜系统的各细胞器固定的毗邻关系,便于实现内膜系统各个部分之间通过膜泡运输的实现物质运输。
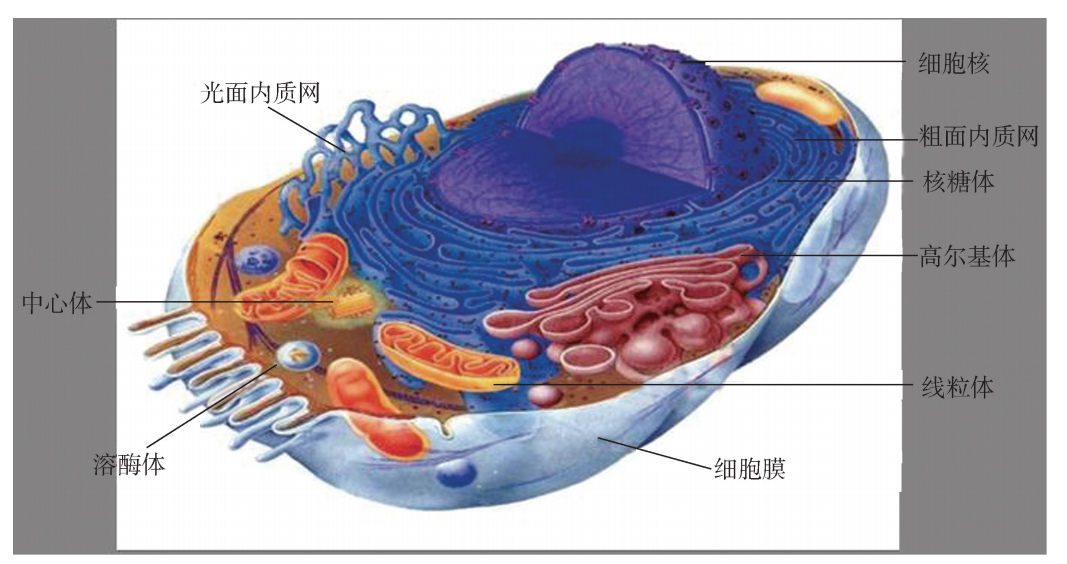
图1 内膜系统
第一节 内质网
1945年,由 K等于1945年通过电镜观察培养的小鼠成纤维细胞时,发现细胞质内部有一些分布在核周围的相互吻合的网状结构,故将其命名为内质网 (endoplasmic reticulum,ER)。见图2 原核细胞和真核细胞的成熟红细胞不含内质网。
图2 内质网电镜图
一、 内质网的化学组成、 结构与分类
(一) 内质网的化学组成
细胞总体积比例( %) | 膜相结构化学成分比例(%) | 脂类和蛋白质比例(%) | 脂类成分 |
| 10 | 50 | 50 | 磷脂、中性脂、缩醛脂和神经节苷脂等;磷脂中磷脂酰胆碱丰富,而鞘磷脂较少。 |
内质网膜的蛋白质和酶类占60%~70%,种类复杂多样,酶类至少有30种,分为5中酶系,具体见下表。
| 解毒有关的氧化反应电子传递酶系 | 细胞色素b5、NADH-细胞色素b5还原酶、NADH-细胞色素c还原酶、 细胞色素p450、NADH-细胞色素p450还原酶 | ||||
| 脂类代谢有关的酶类 | 脂肪酸 CoA 连接酶、磷脂醛磷酸酶、胆固醇羟基化酶、磷脂转位酶 | ||||
| 糖类代谢 有关的酶类 | 葡萄糖 -6-磷酸酶、葡萄糖醛酸转移酶和 GDP-甘露糖转移酶等 | ||||
(二) 内质网的形态结构
一层单位膜构成的内质网膜和内质网腔组成,膜厚5~6nm,形成大小不同的小管、小泡和扁囊状结构相互连接的三维管网结构系统,其网腔相互连通。内质网与外核膜连通,可以延伸到细胞边缘。
(三) 内质网的基本类型
| 类型 | 形态 | 分布 | 应用 |
| 粗面内质网 | 扁囊,表面具有核糖体颗粒 | 蛋白质合成旺盛的细胞:胰腺细胞和合成抗体的浆细胞、分泌细胞、胚胎细胞和肿瘤 细胞 | 肿瘤细胞中粗面内质网的数量与肿瘤细胞的生长率以及恶性程度存在负相 关性 |
| 光面内质网 | 分枝 小管或小泡,无糖体颗粒 | 睾丸间质细胞、卵巢黄体细胞、肝细胞及横纹肌细胞(肌质网) |

图3 内质网结构模式
二、 内质网的功能
(一) 粗面内质网的功能
蛋白质的合成、加工修饰及转运分泌,体现几个 方面:
1 作为核糖体附着的支架
细胞中所有蛋白质的合成,都起始于细胞质中游离的核糖体上,在其合成起始后,必须随核糖体一起转移,附着于粗面内质网上才能最终完成。
2 信号肽介导蛋白质的合成
粗面内质网合成的蛋白质主要有以下四类:
①分泌性蛋白或输出性蛋白,如激素、抗体、酶类、细胞因子以及细胞外基质等;
②膜整合 蛋白,如膜受体、载体蛋白和膜抗原等;
③溶酶体蛋白;
④可溶性驻留蛋白,主要是 指分子量相对较小,可以溶解于水,存在于内质网、高尔基复合体、溶酶体及过氧化 物酶体等膜性细胞器中的蛋白质。
(1)多肽链如何转入内质网
新生的多肽链需要由信号肽介导,与核糖体一起转运移至内质网膜上,并在内质网膜上继续翻译,进入内置网的主要机制就是信号肽假说 (signal hypothesis)
信号肽假说 (signal hypothesis)1971年由德国生物学家Bloble和Dobberstin提出的,认为新生肽链具有一段独特的序列(15~30个氨基酸,称为信号肽,决定蛋白质在胞内的去向或 定位的作用),引导新合成的肽链从细胞质附着于内质网膜上,跨脂双分子层进入内质网腔的,获得了1999年诺贝尔生理学或医学奖。具体过程如下:
信号肽一经合成即可 被细胞质中的信号肽识别颗粒 (signal recognition particle,SRP)受体识别并结合,并 且在移位子 (translocon)的协助下,信号肽引导新生肽链跨脂双分子层进入内质网腔 或直接整合在内质网膜中。移位子也称为转位器 (translocation apparatus),是内质网膜上的一种亲水通道,是一种动态结构,以两种可以转化的构象形式存在。即当它和信号肽结合时,处于一种开放的活性状态;在蛋白质多肽链被完全转移之后,则转变为无活性的关闭状态。
新生肽链进入内质网腔的基本步骤大致如下:第一步,信号肽的识别及其与信号肽识别颗粒的结合:内质网膜上存在信号肽识 别颗粒受体 (SRP-receptor,SRP-R)和移位子,细胞质中存在信号肽识别颗粒。 SRP由6个多肽亚单位和1个 RNA 构成的复合体,具有翻译暂停区域、信号肽识别区 域、受体结合位点,既能被露出核糖体外的信号肽识别、结合,又能识别粗面内质网 上的SRP受体。新生肽链的信号肽一旦被翻译,即被SRP识别并结合,导致翻译暂时 停止,肽链的延长受到阻遏。SRP的另一端与核糖体结合,形成SRP-核糖体复合体, 引导向内质网移动,与内质网膜上的 SRP-R 识别并结合,形成一个 SRP-信号肽- 核糖体-mRNA 复合体锚定在内质网膜通道蛋白移位子上,附着核糖体形成。SRP- R也被称为锚泊蛋白 (dockingprotein),该蛋白能够通过与SRP的识别而使得核糖体 结合附着于内质网上,是内质网的一种膜整合蛋白。当 SRP- 信号肽 - 核糖体 - mRNA 复合体结合在内质网膜上,随即 SRP被解离,返回细胞基质中重复上述过程。 此时,暂时被阻遏的肽链延伸又继续合成 (图4)。同时内质网膜上蛋白质转运通道 打开,核糖体与通道结合,信号肽插入通道。
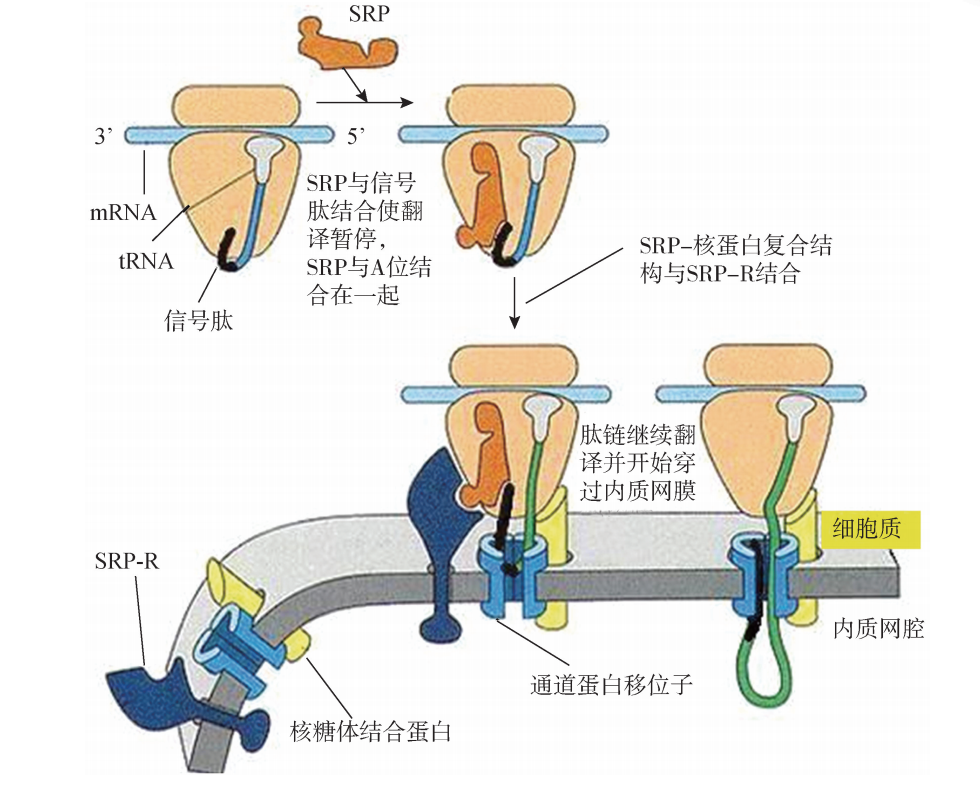
图4 信号识别颗粒作用过程
第二步,新生肽链进入内质网腔:在信号肽的引导下,合成中的肽链通过核糖体 大亚基的中央管和移位子蛋白共同形成的通道,穿过内质网膜进入内质网腔。随之, 信号肽序列被内质网膜腔面上的信号肽酶切除,新生肽链继续延伸,直至完成而终止。 随后,完成肽链合成的核糖体大、小亚基解聚,与内质网分离。脱离内质网的核糖体 回到细胞质,进行核糖体的再循环。
(2) 蛋白质的折叠与装配:
多肽链合成之后,需要按照特定的方式折叠形成三维 空间结构,内质网为新生多肽链的正确折叠和装配提供了有利的环境。多肽链的折叠过程受分子伴侣 (molecular chaperone)的调节,分子伴侣是一类在细胞内能够帮助其 他多肽链进行正确折叠、组装、转运的蛋白质。分子伴侣属于热激蛋白家族,能够识 别并滞留那些折叠组装错误的蛋白质而不被运输,也能够抑制未成熟蛋白质的折叠并 使其降解。
分子伴侣不参与最终产物的形成,如内质网标志性分子伴侣,葡萄糖调节蛋白94。内质网腔中的氧化型谷胱甘肽 (GSSG)和内质网膜腔面上的蛋白二硫键异构 酶 (PDI),为二硫键的形成及多肽链快速折叠提供了保证,这个过程需要分子伴侣的帮助。分子伴侣对新生蛋白质的折叠,通常始于多肽链的某些区域。分子伴侣蛋白质 的共同特点是在羧基端有一个 Lys-Asp-Glu-Leu四氨基酸滞留信号肽,它们能够 和内质网膜上的相应受体结合而驻留于网腔不被转运,因此也被称为驻留蛋白 (retentionprotein)。
(3)蛋白质的糖基化
糖基化 (glycosylation)是指多肽链中特殊的氨基酸残基侧链上,以共价键连接上单糖和低聚糖形成糖蛋白的过程。由附着核糖体合成并经内质 网转运的蛋白质,大多数都要被糖基化。
糖基化分2种类型:
N-连接糖基化,糖链被连接在多肽链中天冬酰胺 (Asn)残基的-NH2 端,发生在粗面内质网腔内。N-连接糖蛋白的糖基化主要有两类:第一类是高甘露糖,主要由 N-乙酰葡萄糖胺 (2分子)、葡萄糖 (3分子)和多个甘露糖 (>9分子)组成;第二类是复合寡糖, 主要由 N-乙酰葡萄糖胺、半乳糖、唾液酸以及岩藻糖组成。

O-连接糖基化,单糖链被连接在多肽链中的丝氨酸 (Ser)或 苏氨酸 (Thr)残基侧链的-OH 端上,发生在高尔基体腔内。
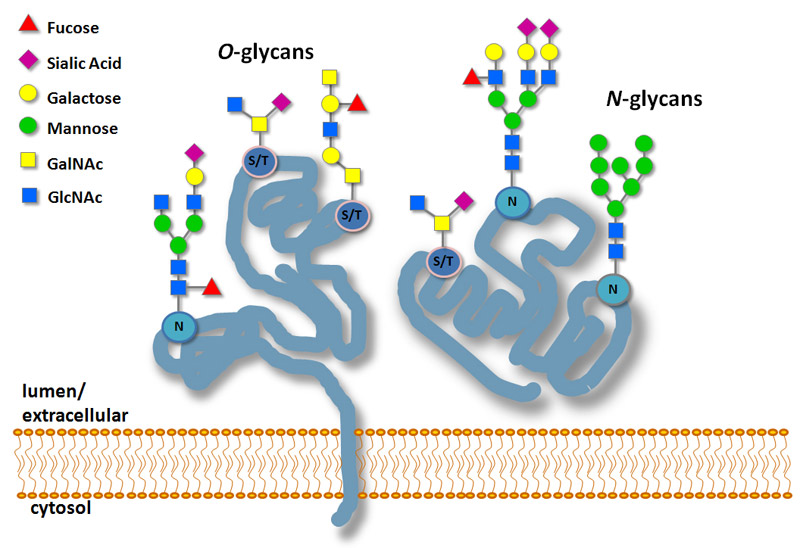
糖基化修饰的生物学意义:
①糖基化使蛋白质免遭水解酶的降解,因而对蛋白质具有保护作用;
②糖基化能够引导蛋白质包装形成运输小泡,使蛋白质进行靶向运输,因而糖基化对蛋白质具有运输信号的作用;
③糖基化在细胞膜表面形成糖被,在细胞膜的保护、识别以及通讯联络等生命活动中发挥重要作用。
(4)蛋白质的胞内运输
经过粗面内质网的修饰和初步加工之后外输蛋白,被内质网包裹形成膜性小泡,以出芽的方式与内质网分离开来,通过两种途径被进一步运输:
①是与高尔基体 形成面的小囊泡融合,并在高尔基体腔内浓缩加工为分泌颗粒,脱离高尔基体,以胞吐形式分泌到细胞外,如抗体和消化酶等。 ②一种途径是这种小泡直接与细胞质中 的浓缩泡融合,形成酶原颗粒,然后被排出细胞,如溶酶体蛋白。
(二) 光面内质网的功能
1 参与脂类的合成与转运
生物膜中的磷脂、胆固醇和糖脂均在光面内质网合成。内质网合成脂质的底物来自细胞质基质,合成脂质所需的相关酶类定位于内质网膜上的镶嵌蛋 白,但催化作用是在内质网的细胞质侧完成的。
主要过程:首先是磷脂酸的形成,由脂酰基转移酶催化脂酰辅酶 A 的两条脂肪酸链转移并结合到甘油G-3-G磷酸分子上生成;其次是双酰基甘油的形成,是由磷酸酶催化磷脂酸去磷酸化而生成;最后在胆碱磷酸转移酶的催化下,在双酰基甘油上添加结合一个极性基团,最终形成由一个极性头部 基团和两条脂肪酸链疏水尾部构成的双亲性脂类分子。
光面内质网中合成的脂质分子在翻转酶的作用下,快速由细胞质基质侧转向内质网腔面,其中一部分嵌入到内质网膜的脂类双层中,另外一部分以出芽的方式转运至高尔基体、溶酶体等其他膜性细胞器以及细胞膜。
2 参与糖原的合成与分解
肝脏和肌肉是糖原储存最主要的组织器官,在这些组织中光面内质网较为发达,大量的糖原颗粒靠近光面内质网。实验证明,肝细胞中的光面内质网膜上含有葡萄糖-6-磷酸酶,能够将其表面的糖原降解为葡萄糖-6-磷酸酶,使之去磷酸化,形成葡萄糖和磷酸,然后将葡萄糖注入内质网腔中,经过内质网膜运输到血液中补充血糖。
3 解毒作用
肝脏是机体中内、外源性毒物和药物分解解毒的主要器官,肝脏的解毒作用主要由肝细胞中的光面内质网完成。光面内质网上集中了多种重要的氧化及 电子传递酶系,如细胞色素 P450、NADPH- 细胞色素 P450 还原酶、细胞色素 b5、 NADPH-细胞色素c还原酶等。
光面内质网解毒机制包括三个方面:
①氧化和羟化作 用,即药物或者毒物经光面内质网膜上的氧化或羟化后,消除其作用和毒性,也许这 种氧化作用可能会使某些物质的毒性增强,同时由于羟化使代谢产物极性增强而易于 排出体外;
②转化作用,如许多氨基酸代谢生成的氨可以转化成无毒的尿素,经肾脏 排出体外;
③结合作用,某些药物可以结合葡萄糖醛酸,形成水溶性物质,从而易于 排出体外。
4 参与甾体类激素的合成
在生殖腺和肾上腺的内分泌细胞中,光面内质网和线 粒体,可能还有高尔基体上的一些酶共同参与甾体类激素的合成。
5 其他功能
光面内质网在特化的细胞中执行特殊功能。
①调节肌肉收缩 运动,骨骼肌细胞中的光面内质网特化为肌质网,肌质网具有储存和调节 Ca2+ 浓度的 功能,肌质网通过其膜上的Ca2+ 通道和Ca2+ 泵调节细胞内的Ca2+ 浓度,从而调控肌肉 的收缩和舒张;
②分泌盐酸和调节渗透压,胃底壁细胞的光面内质网具有分泌盐酸和 调节渗透压的功能。

