
第四节 细胞膜的信号转导功能
生物细胞每时每刻都在接受来自细胞内外各种各样的信号,信号转导是生命活动的一种最基本和最重要的方式。各种信号调控着细胞的一切生命活动,引起机体各组 织器官的发育与衰老。细胞对胞外信号的反应就是细胞与体内外环境及细胞之间电信号、化学信号等的传递、识别及应答的过程。
一、 信号转导的意义
(一) 信号转导概述
信号转导 (Signaltrans duction)是化学或物理信号通过与细胞膜上或胞内的受体 特异性结合,将信号转换后传给胞内相应的系统,最终导致细胞对信号做出适当的反 应。负责接收信号分子的蛋白质被称为受体。信号通路能够相互作用,形成信号网络, 协调细胞内部反应。在分子水平上,这样的反应包括基因转录或翻译的改变,以及翻译后的顺应性;还包括蛋白质含量和活性的变化,以及它们在位置上的变化。这些分子事件是控制细胞生长、增殖、代谢等许多过程的基本机制。在多细胞生物中,信号转导途径已进化为以多种方式进行细胞的通讯,从而调控机体的各种生理反应,伴随人体发育的一生。
(二) 信号转导的临床意义 正常有序的信号转导是细胞功能与代谢的基础,信号转导异常可以导致各种病理过程。信号转导和疾病关系的研究是当前生命科学研究的一个热点,随着研究的不断深入,不仅阐明了多种遗传疾病的发生机制,而且证实了许多危重病,如炎症、感染、 脑血管疾病、糖尿病、恶性肿瘤等的发病与信号转导异常有着密切的关系。深入研究信号转导对于探讨疾病的发生、发展和防治具有重要的临床意义。设计以细胞信号转 导通路为靶点的药物和疾病治疗方法,也已经成为临床医学和药物产业的新领域。由于信号转导的复杂性,应用系统化和网络化的观念与方法进行深入的研究,才有可能 具有实用的临床意义。
二、 信号转导途径
(一) 信号转导途径的基本组成
1 细胞间通讯的方式
细胞间通讯 (Intercellular communication)是指细胞与细 胞之间的通讯,有四种类型 (图1)。(1)内分泌信号传递 (endocrine signaling):内分泌细胞 (endocrine cell)作为整 体控制细胞行为的第二种特化的信号细胞,分泌激素信号分子到血流中,血流再将信号运载到身体内广泛分布的靶细胞。(2)旁分泌信号传递 (paracrine signaling):大多数情况下信号分子是旁分泌信 号,对邻近的细胞起调控作用。旁分泌信号迅速地被周围靶细胞吸收,被胞外酶降解, 或者被胞外基质固定。(3)突触信号转导 (synaptic signaling):当来自环境或其他神经细胞的信号激活 神经细胞时,神经细胞沿着其轴突迅速地传递电脉冲,当该脉冲到达轴突末端时,导 致那里的神经末梢分泌称为神经递质 (neurotransmitter)的化学信号,化学突触的结 构能确保神经递质特异地传递到突触后的靶细胞。(4)接触依赖的信号传递 (contact-dependent signaling):某些信号分子结合在 信号细胞的表面上,只影响与之接触的细胞,在发育和免疫应答中尤其重要。
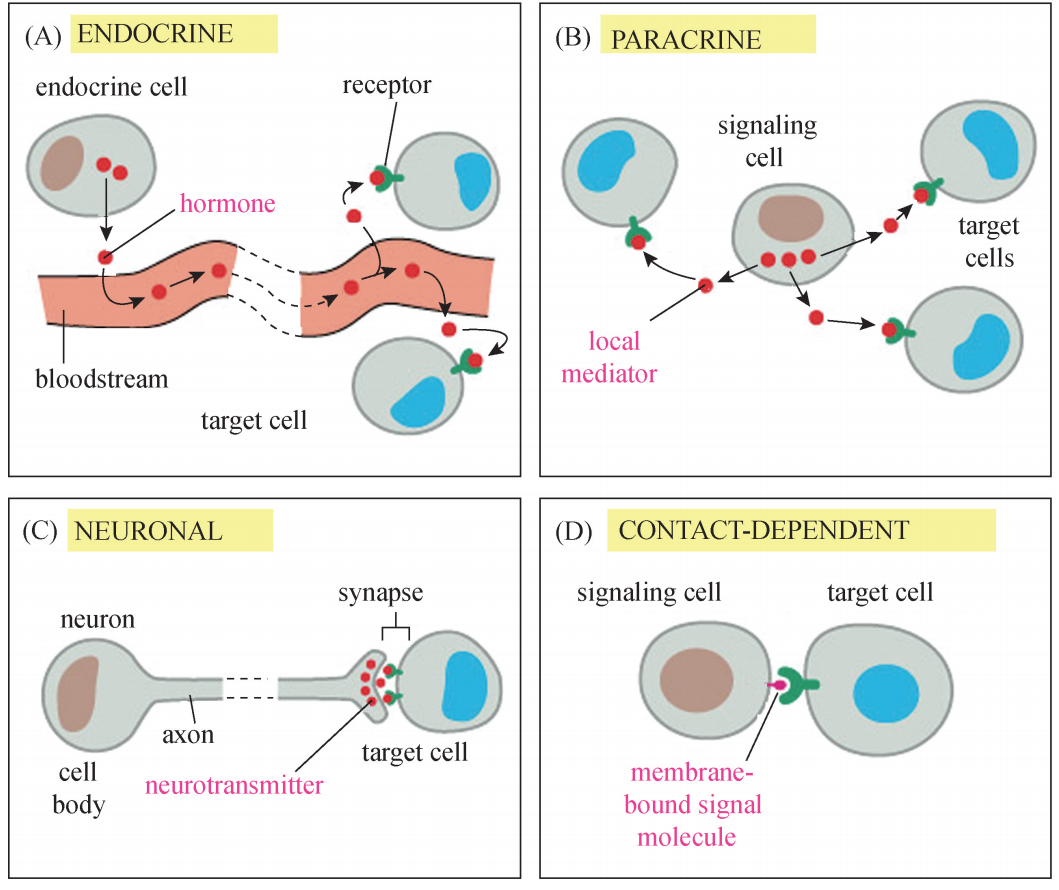
图1 细胞间通讯示意
2 细胞信号转导的基本组成 四种类型的细胞通讯都包括以下4个部分 (图2):
(1)胞外信号分子,包括激素、神经递质、细胞因子、气体分子等,通常也称为第一信使 (firstmessenger)。这些信号分子,从化学结构来看,他们主要包括肽类分 子、氨基酸、脂类、胆固醇衍生物、气体分子 (NO、CO2)。它们都具有特异性、高 效性和可被灭活的特点,只能与特定的受体结合,几个分子就可以产生明显的生物学 效应,完成信息传递后可以被降解或修饰失去活性。
(2)细胞表面受体和胞内受体:细胞表面或内部存在能够接受胞外信号分子的受 体,与相应的信号分子特异地结合从而实现对信号的定向转导。脂溶性小分子信号分 子能够进入细胞与胞内受体结合发挥作用,这种受体称为胞内受体或核受体,但大多 数信号分子本身并不进入细胞,需要通过细胞表面的受体及相关分子将信号传入,这 类受体称为膜受体。常见的膜受体有:①生长因子类受体,这类受体具有酪氨酸激酶 或者丝氨酸/苏氨酸激酶的活性,能直接催化底物的磷酸化;②神经递质的受体,其本 身是一种或几种离子的离子通道。配体与这类受体结合后,改变了受体的空间构象, 使离子通道开放或关闭,控制着离子进出细胞;③G 蛋白偶联的受体,是神经递质、 激素、肽类和胺类配体的受体。
(3)胞内信号分子:膜受体将信号分子所携带的信号传递到细胞内,其中部分受 体在进行细胞信号的跨膜传递过程中产生如cAMP、cGMP、Ca2+ 等胞内小分子信号, 称为第二信使 (secondmessenger)。
(4)胞内效应分子:胞内第二信使分子通过细胞内的一系列下游分子进一步传递 信号,最终作用于胞内的效应分子,使其进入细胞核启动基区转录,导致细胞各种复 杂的生物学效应。胞内受体自身通常就是转录因子,与进入细胞内的信号分子结合后 改变构象进入细胞核,调节基因转录。
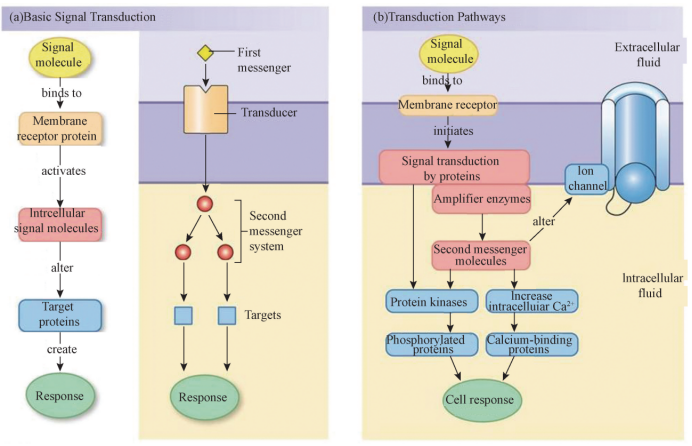
(二) 胞内信号传递途径 细胞内信号传递径经种类众多,这里重点介绍两种。
1G蛋白偶联受体信号转导途径 (1)G 蛋白的概况:G 蛋白 (GProtein)的全称为鸟苷酸结合蛋白 (guanine nucleotide bindingprotein),一般是指任何可与鸟苷酸结合的蛋白质的总称 (图)。 G 蛋白的特征是:①由α、β、γ等3个不同的亚单位构成的异聚体;②具有结合 GTP 或 GDP的能力,并具有 GTP酶 (GTPase)的活性,能将与之结合的 GTP 分解形成 GDP;③其本身的构象改变可进一步激活效应蛋白 (effectorprotein),使后者活化, 实现把细胞外的信号传递到细胞内的目的。G 蛋白分为3类:Gs、Gi和 Gq。其区别是 α亚基结构和活性的不同:Gs指 G 蛋白的a亚单位对效应蛋白起激活作用;Gi指 G 蛋 白的a亚单位对效应蛋白起抑制作用;Gq其功能尚未清楚。
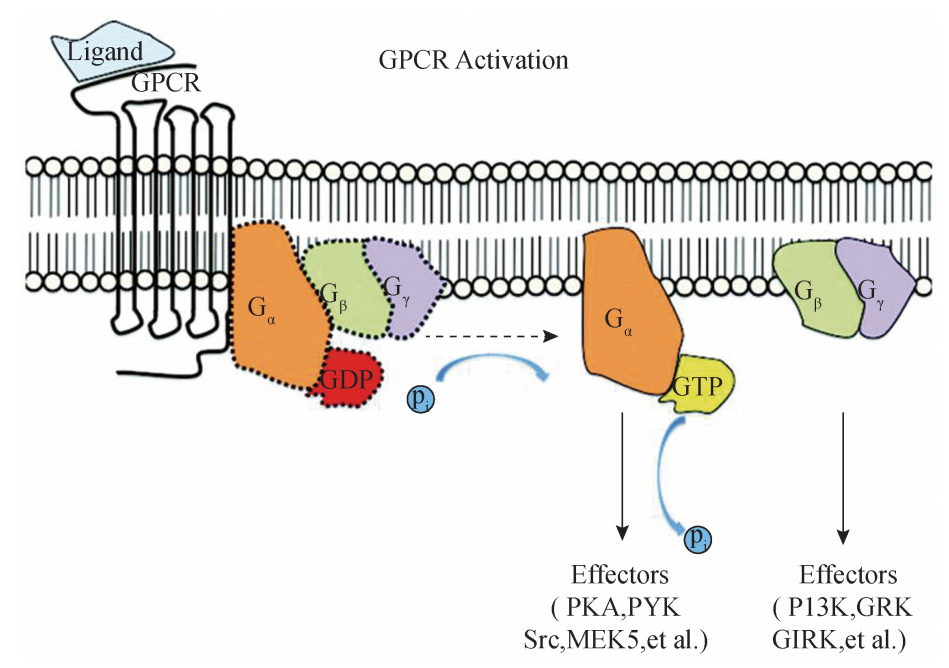
(2)G 蛋白偶联受体信号转导通路:G 蛋白偶联受体介导的信号转导是最典型的 膜受体信号途径,环境中的激素、神经递质、趋化因子和光线等各种因素能激活 G 蛋 白偶联受体,并触发细胞内的一系列信号通路.当细胞膜外信号分子与 G 蛋白偶联受 体结合后,受体构象变化,进一步激活附着在其细胞膜内侧端的异三聚体 G 蛋白,表 现为 G 蛋白上原先结合的 GDP被替换为 GTP;激活后的 G 蛋白会进一步激活其效应 蛋白,引发一系列的下游效应,其中所涉及的具体信号通路则取决于 G 蛋白的种类. 下面重点介绍两种类型. 1)腺苷酸环化酶与 CAMP信号转导通路:激素、神经递质等第一信使与相应的 膜受体结合后,可以激活 G 蛋白,并活化位于细胞膜上的 G 蛋白效应蛋白———腺苷酸 环化酶 (adenylate cyclase,AC),使 ATP转化生成第一信使cAMP,进一步分别引起 相应底物的磷酸化级联反应、离子通道活化等效应,参与调节细胞代谢、增殖、分化 等不同生 理 过 程.在 绝 大 多 数 细 胞,cAMP 特 异 地 活 化 cAMP 依 赖 性 蛋 白 激 酶 A (cAMP dependent protein kinaseA,PKA)来调节细胞的新陈代谢.一般而言,PKA可使某些 特 殊 的 底 物 蛋 白 磷 酸 化,这 种 底 物 蛋 白 通 常 是 cAMP 反 应 元 件 结 合 蛋 白 (cAM Presponsive element binding
proteinC,CREB)等基因表达的调节因子,激活后 的 CREB可结合相关基因的 CRE 区 (序列为 TGACGTCA),在其他特异性转录因子 的协助下,启动基因的表达,表达的蛋白质产物对细胞产生各种生物学效应 (图3G4G 4).在不同的组织中,依赖cAMP的蛋白激酶的底物大不相同,cAMP通过活化或抑 制不同的酶系统,使细胞对外界不同的信号产生不同的反应 (图4)。
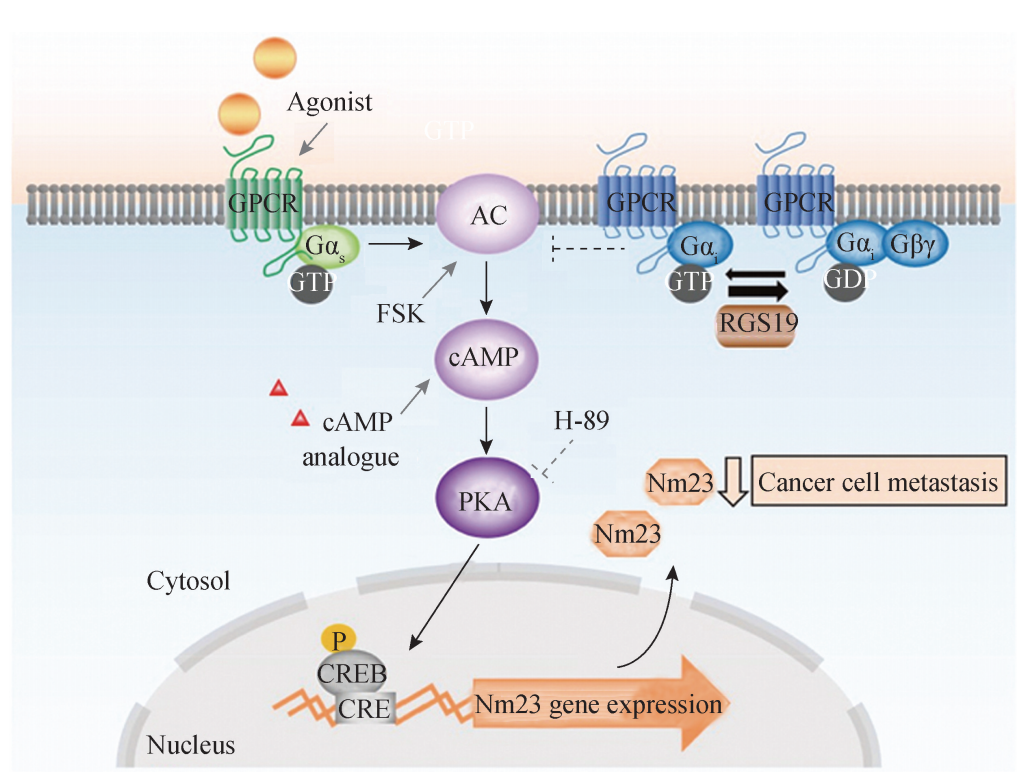
腺苷酸环化酶与cAMP信号转导通路
2)磷脂酰肌醇信号通路:神经递质如毒蕈型乙酰胆碱、α1 肾上腺素、5-羟色胺 等,多肽激素如v1-后叶加压素、血管紧张素 H、p物质和促甲状腺素释放因子等, 生长因子如血小板生长因子 (PDGF)、T 细胞有丝分裂原 (植物凝集素和刀豆球蛋白 A)等,它们作为胞外信号与膜受体结合,激活膜上的 G 蛋白活化磷脂酶 C (phospholipase C,PLC)催化细胞膜上的4,5-二磷酸酯酰肌醇 (phosphatidylinositol4,5- biphosphate,PIP2)分解为两个重要的细胞内第二信使———DAG 和IP3。 脂溶性 DAG 增多后,蛋白激酶 C (protem kinaseC,PKC)紧密结合在膜的内 面,与 DAG 结合被活化,激活后 PKC对 Ca2+ 的亲和力增强,从而实现其对底物蛋白 酶的磷酸化功能,如催化与分泌及增殖有关的蛋白磷酸化,还能活化 Na+ -H+ 交换系 统,使细胞内 H+ 减少,提高细胞质中的pH 值,还可以增强 Na+ -K+ 泵的运转等。 IP3 为水溶性物质,它从细胞膜扩散到细胞胞质,与内质网膜上的IP3 受体结合, 调控打开肌浆 (内质网)膜上的 Ca2+ 通道,Ca2+ 由内质网腔释放入细胞质,提高细胞 质中游离 Ca2+ 的浓度。Ca2+ 与钙结合蛋白钙调素 (calmodulin,CAM)结合后,引起 构象改变,形成 Ca2+/CAM 复合物而被活化,活化后可激活蛋白激酶或磷酸酶,后两者可磷酸化底物蛋白,调节细胞内代谢活动 (图5)。
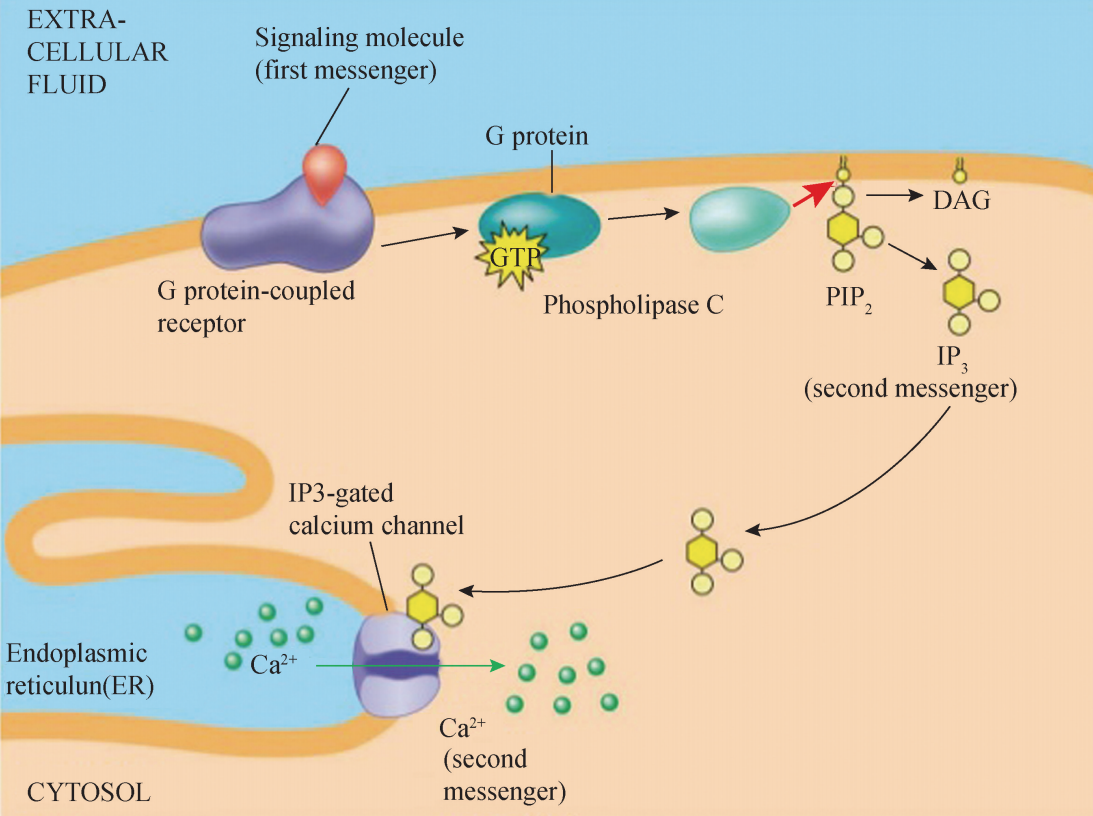
磷脂酰肌醇信号通路
2 受体酪氨酸激酶介导的 RTK-RAS信号途径 受体酪氨酸激酶 (receptortyrosinekinase,RTK)是细胞膜表面常见的多肽生长因子类受体,包括表皮生长因子受 体、血小板生长因子受体、神经生长因子受体等,其结构的共同特点是整个分子可以 分成3个结构区,即细胞外的配体结合区、细胞内部具有酪氨酸蛋白激酶活性的区域 和连接这两个区域的跨膜结构。 受体酪氨酸激酶介导的 RTK-RAS信号通路与细胞增殖、细胞应激、细胞凋亡等 密切相关,具体过程如下:①生长因子 (配体)与 RTK 胞外结构域结合后,首先触发 膜上相邻的 RTK 相互靠近形成二聚体乃至寡聚体,并促进 RTK 胞内区的相互作用, 使胞内段互为对方激酶的底物,引发二聚体或寡聚体的自身磷酸化;②RTK 磷酸化激 活后与 Grb2通过SH2结构域结合,再与Sos的SH3结构结合,Sos激活 Ras(一种定 位于膜上的小分子 G 蛋白);③由于 Ras本身的 GTP酶活性不强,需要 GTP酶活化蛋 白 (GAP)的参与,活化的 Ras蛋白激活丝氨酸/苏氨酸蛋白激酶 (MAPKKK);④活 化的 MAPKKK 结合并磷酸化另一种蛋白激酶 (MAPKK),使其活化;⑤ 活化的 MAPKK 使 MAPK 的苏氨酸和酪氨酸残基磷酸化并使之激活;⑥活化的 MAPK 进入 细胞核,可以使许多转录因子活化,产生效应。Ras-MAPK 途径是多条信号通路共 有的通路,包括多种生长因子、细胞因子、G 蛋白偶联受体、整合素等都与此通路相关 (图6)。
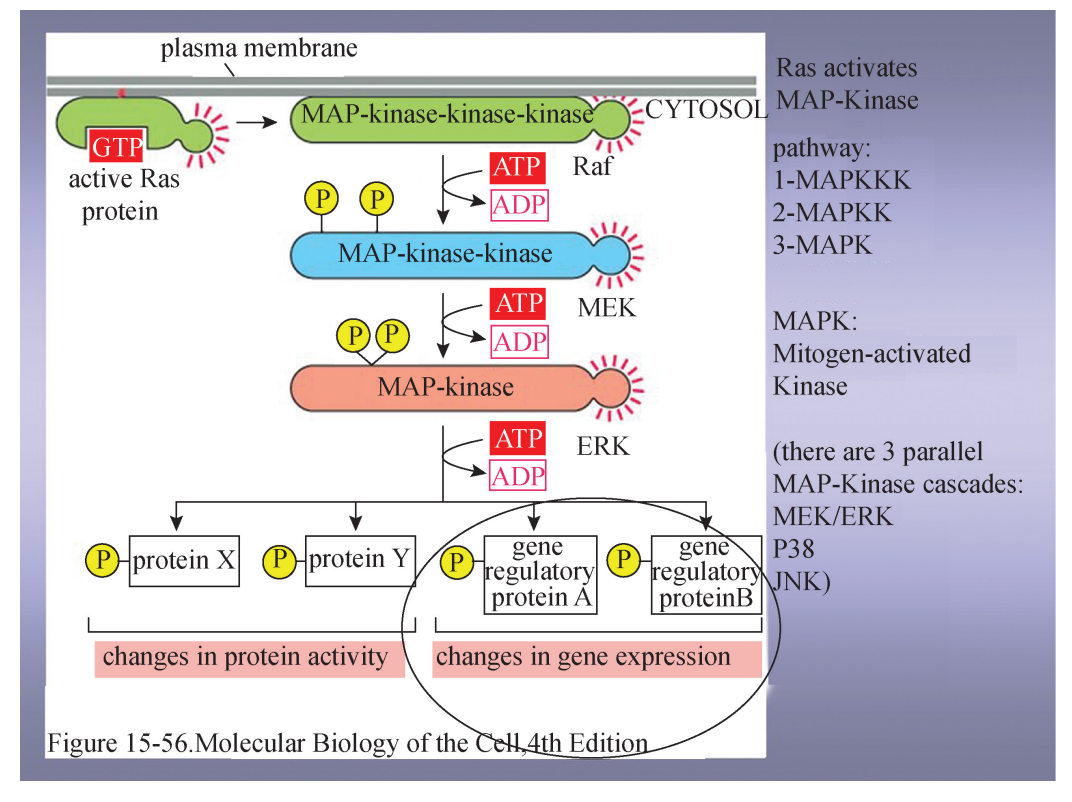
RTKGRAS介导的 MAPK 通路
三、 信号转导异常与临床疾病关系的研究进展 细胞信号转导异常会导致疾病,常常涉及受体、胞内信号转导分子及信号转导通 路等多个环节。
1 受体病 因受体的数量、结构或调节功能变化,使之不能介导配体在靶细胞中 应有的效应所引起的疾病。如家族性高胆固醇血症 (familial hypercholesterolemia,FH),为常染色体显性遗传病,是由于基因突变引起的LDL受体缺陷症,导致LDL受 体数量减少,导致胆固醇在心血管积累,伴发冠心病、动脉粥样硬化等症。按 LDL受 体突变的类型及分子机制可分为:受体合成障碍、受体转运障碍、受体与配体结合障 碍、受体内吞缺陷、受体再循环障碍等。重症肌无力 (myadstheniagravis)是一种主 要累及神经肌肉接头突触后膜上乙酰胆碱受体的自身免疫性疾病,患者的体内产生了 抗乙酰胆碱受体的抗体,抗体与乙酰胆碱受体结合,封闭了乙酰胆碱的作用,并促进 乙酰胆碱受体的分解,使患者体内受体的数目明显减少,使通过乙酰胆碱受体进行的 信号转导过程障碍,而出现重症肌无力病症。其他还有 VitD 抵抗性佝偻病、甲状腺素 抵抗综合征、雌激素抵抗综合征等。
2G 蛋白功能异常 霍乱是由于霍乱弧菌附于小肠黏膜进行繁殖而引起的急性腹 泻。由霍乱弧菌所产生的霍乱毒素由α、β两个亚基组成,其中β亚基可与细胞膜上的 受体结合,α亚基能穿过细胞膜,催化细胞内的 NAD+ 中的 ADP核糖基不可逆地结合 在 Gs的α亚基上,使α亚基和β、γ亚基分离并与 GTP 结合,但此时,亚基丧失了 GTP酶的活力,因而不能把 GTP水解为 GDP,所以 G 蛋白处于持续激活状态,同时AC被活化了的α亚基持续激活,从而使细胞中的cAMP大量增加,可高达其正常值的 100倍以上,促使大量的 Cl- 和 HCO3- 离子从细胞内进入肠腔,细胞内外渗透压失去 平衡,引起大量水分进入肠腔,造成剧烈的腹泻。
3 肿瘤 绝大多数的癌基因表达产物都是细胞信号转导系统的组成部分,可以从 多个环节干扰细胞信号转导过程,导致肿瘤细胞增殖和分化异常,主要有以下两个方 面:(1)促进细胞增殖的信号转导过强:①某些癌基因可以编码生长因子样的活性物 质,如sis编码PDGFβ链同源物,intA 编码FGF类似物;②受体过度表达或原癌基因 表达 GF受体类蛋白质,如erbGB编码变异型 EGF受体;③通过编码非受体 TPK 或丝 /苏氨酸激酶影响信号转导,如src编码 TPK,raf编码Ser/ThrKinase;④ras编码小 分子 G 蛋白,在肿瘤组织中ras突变 (甘氨酸12、甘氨酸13、谷氨酸61 为其他氨基酸取代),变异的ras与 GDP解离升高,GTP酶活性降低,可以使ras持续活化,cAMP升 高导致细胞增殖;⑤c-myc、c-fos、c-jun、Ets表达产物位于核内,能与 DNA 结合调 节转导活性,使癌基因超表达。(2)抑制细胞增殖的信号转导过弱,主要是由于生长 抑制因子受体的减少、丧失或信号转导通路异常所致。
思考题
1 细胞膜的特性有哪些?
2 简述葡萄糖转运的过程?
3 简述 LDL颗粒的转运过程?
4 细胞信号转导基本途径有哪些?

