-
1 一、跨膜运输
-
2 二、 膜泡运输
-
3 三、 膜运输...
第三节 细胞膜的物质运输功能
学习要求
知识内容
细胞与细胞、细胞与基质、细胞与外环境之间都需要进行物质运输,既要把细胞所需要的物质 (如营养物质)不断摄入细胞内;同时也要把细胞新陈代谢的产物及时地分泌或排出细胞外。进出细胞的物质种类繁多,分子量大小相差悬殊,进出细胞的 方式也有很大的不同。由于膜具有流动性,即磷脂分子和膜蛋白分子之间的距离是可在一定范围内变化的,因此分子量小的粒子可以穿过细胞膜进行运输,而分子量大的粒子则无法穿过细胞膜,必须采取膜泡运输的方式进出细胞。
一、跨膜运输
跨膜运输是离子和小分子进出细胞膜的方式。由于构成细胞膜的脂双层的中间部分是疏水的,所以细胞膜是选择半透性屏障,只有小分子和非极性的物质可以自由出入细胞膜,绝大多数极性小分子和离子是不能自由透过细胞膜的,必须以被协助的方式通过细胞膜。离子和小分子的跨膜运输可分为被动运输(passivetransport)和主动运输(activetransport)两大类。
1被动运输被动运输是指物质顺浓度梯度,从浓度高的一侧经细胞膜转向浓度低的一侧。被动运输又可分为简单扩散(simple diffusion)、渗透(osmosis)和促进扩散(facilitated diffusion)。
(1)简单扩散:简单扩散是分子从较高浓度到较低浓度的运输或移动而不消耗能量。在这个过程中,分子只是通过细胞膜的孔扩散。简单扩散不需要转运蛋白。当细胞膜内部和外部的分子浓度不同时,建立浓度梯度,所含的势能(potential energy),然后分子从较高浓度移动到较低浓度,直到保持平衡。当膜的两侧分子浓度变得相等时,运输过程停止。扩散速率由细胞质膜的浓度梯度和渗透性决定。细胞膜的浓度梯度和渗透性越大,被动扩散的速度就越大。可以借助单纯扩散通过细胞膜的物质只有两类,第一类是疏水的(脂溶性的)小分子,如氧、氮、苯等,其中脂溶性越大的扩散越快;第二类是不带电的极性小分子,如水、二氧化碳、乙醇、尿素、甘油等,其中分子量越大的扩散速度越慢(图1)。所以像葡萄糖这类不带电的极性分子因分子量太大,几乎不能自由扩散过膜。水能很快通过脂质双分子层,这是由于水分子很小,不带电荷,且有双极结构,因而能迅速地通过脂质双分子层。

简单扩散
(2)渗透:
渗透是被动运输的特殊情况,是指水从低渗 (低溶质浓度)溶液扩散到高渗 (高溶质浓度)溶液。一般来说,水流的方向是由溶质浓度决定的,而不是溶质分子本身的性质决定的。例如,在不同浓度的盐水溶液 (高渗、等渗和低渗)中的血细胞出现不同的变化。高渗意味着盐水溶液含有比血细胞更高浓度的溶质,流体将从低溶质浓度 (血细胞)的区域流动到高溶质浓度 (水溶液)的区域。结果使血细胞收缩。如果盐水溶液是等渗的,它将含有与血细胞相同的溶质浓度。流体将在血液细胞和水溶液之间均匀流动。结果,血细胞将保持相同的大小。与高渗溶液相反,低渗溶液意味着盐水溶液含有比血细胞低的溶质浓度和更高的水浓度。流体将从低溶质浓度 (水溶液)的区域流动到高溶质浓度的区域 (血细胞)。结果,血细胞会膨胀甚至破裂 (图2)。
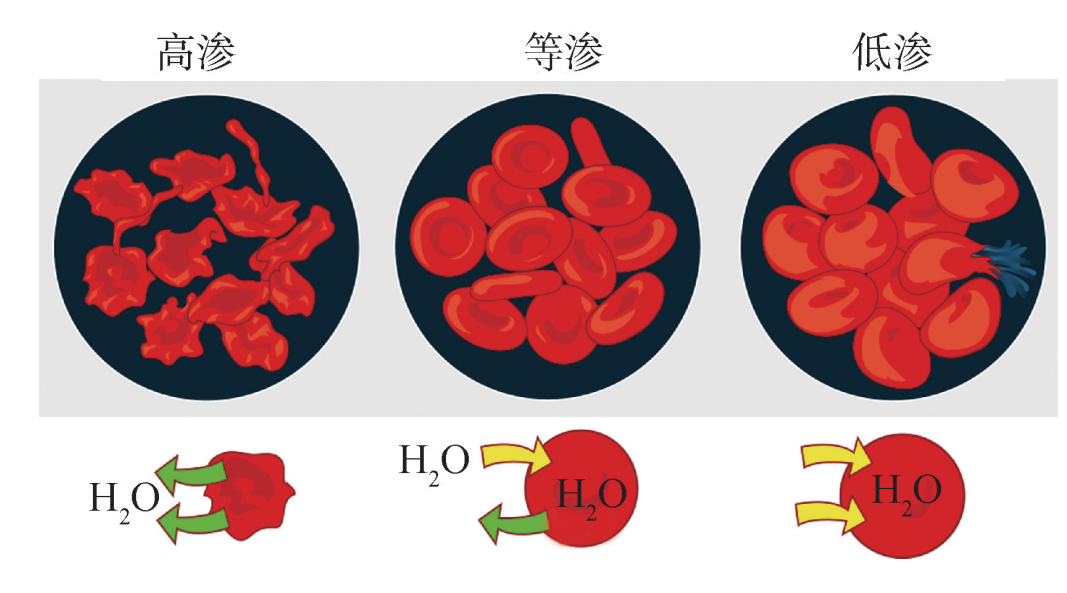
图2 红细胞在不同浓度溶液中的形态
(3)促进扩散:促进扩散是一种被动运输,是一些物质在特殊转运蛋白的帮助下顺着浓 度 梯 度 的 跨 膜 转 运 方 式。 特 殊 转 运 蛋 白 主 要 有 两 类: 通 道 蛋 白 (channel protein)和载体蛋白 (carrier protein)。一些分子和离子如葡萄糖、钠离子和氯离子不能通过细胞膜的磷脂双层,必须通过嵌入在细胞膜伤上的通道蛋白 (channel protein) 或载体蛋白 (carrier protein),把这些物质运输到细胞中。通道蛋白允许特定的离子通 过蛋白质通道,称为离子通道 (lon channel)。载体蛋白与特定的被运输分子结合后, 改变自身形状,将分子从细胞膜外转运到细胞膜内,转运完成后,蛋白质就回到原来 的状态。如葡萄糖、氨基酸、核苷酸和许多离子,必须借助专一的载体蛋白来完成转运 (图3)。
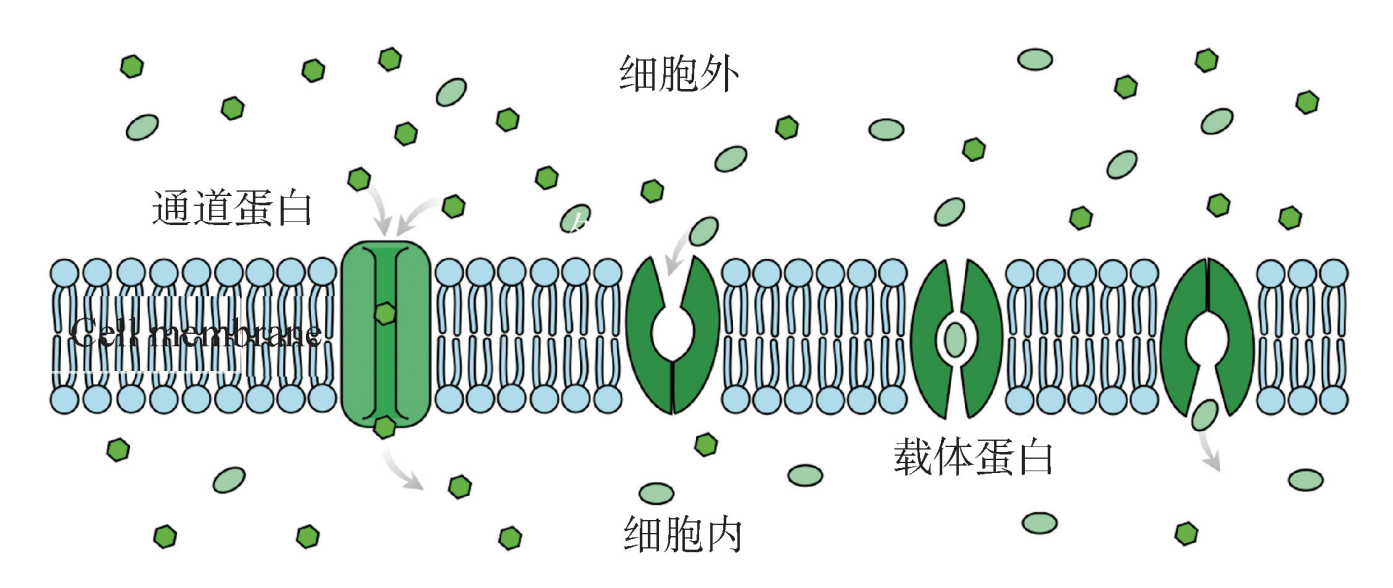
图3 通道蛋白和载体蛋白
离子通道是一种贯穿膜全层的膜蛋白,它们在膜上形成许多直径为0的小孔,允许离子穿过通道孔。它们的功能包括建立静息膜电位、形成动作电位和其他电信号,通过控制离子穿过细胞膜的流动,控制跨分泌细胞和上皮细胞的离子流动,并调节细胞体积。离子通道存在于所有可兴奋细胞的膜中,按照其调控方式的不同分为非门控离子通道 (nongated ion channel)和门控离子通道 (gated ion channel)。
非门控离子通道也称为泄漏离子通道 (leak ion channel),是一种完整的跨膜蛋白,始终开放,并允许一 个 或 多 个 离 子 沿 与 它 们 的 浓 度 和 电 荷 梯 度 一 致 的 方 向 扩散。例如,Duchenne型肌营养不良,是由于 患 者 缺 少 将 细 胞 骨 架 连 接 到 细 胞 外 基 质 的 蛋 白 质, 从 而 导 致 患 者 肌 肉 出 现 高 频 率 的 肌 膜 破 裂, 导 致 钙 泄 漏 通 道(calcium leak channels)过度表达,大量的钙 离 子 进 入 细 胞, 从 而 导 致 肌 肉 细 胞 内钙稳态消失最终导致细胞死亡。
门控离子通道是一种完整的跨膜蛋白,是响应于某些刺激而打开或关闭的离子通道。刺激门控离子通道的因素有膜电位 (电压)变化、特定配体或信号分子的到达和结合 (激素、神经递质)、局部荷尔蒙或机械压力或光能等。
根据刺激信号的性质可将门控离子通道分为:①电压门控离子通道 (voltagegated ion channel)是位于细胞膜外侧的离子通道,当细胞膜电位 (电压)的变化,即局部膜电荷从负到正的变化,可引起离子通道打开或关闭的离子通道。这些通道主要负责去极化事件,即神经冲动在神经元、肌细胞和腺细胞等细胞的传播传导;②化学门控离子通道chemically gated ion channel),可以与特定配体或信号分子 (激素、神经递质、局部激素)结合,打开和关闭离子通道,如神经元、肌细胞和腺细胞的初始去极化;③机械门控离子通道 (mechanically gatedion channel)位于细胞膜外侧,当细胞受到机械压力或振动的刺激, 可以打开和关闭该类通道,如感觉细胞对触摸、振动、压缩或拉伸的响应;④光门控离子通道 (light gatedion channel),受光能的刺激,调节通道的开放或关闭,如感光细胞 (视杆细胞、视锥细胞)。
2主动运输
主动运输 (active transport)是指物质从低浓度的一侧通过细胞膜向高浓度一侧的转运。由于运输是逆着浓度梯度进行的,需要载体蛋白的参与,且消耗代谢能,这类运输方式称为主动运输。例如,在生理条件下,人红细胞内的 K+ 浓度为血浆中的30倍,而细胞内的 Na+ 浓度比细胞外低13倍,但 K+ 仍可由血浆进入细胞内,Na+ 仍可由细胞内转到血浆中,这就需要细胞通过消耗能量、逆浓度梯度主动运输 K+ 和 Na+ 来实现。主动运输可分为初级主动运输 (primary activetransport)和次级主动运输 (secondary active transport)两类。
(1)初级主动运输:初级主动运输直接使用化学能量源 (如 ATP)来移动分子穿过膜以对抗其梯度,比较有代表性的如 Na+/K+ -ATPase酶,又 叫 Na+ -K+ 泵。
Na+/K+ -ATPase酶也叫钠钾腺苷三磷酸酶,存在于所有动物的细胞膜上。具有载体和酶的双重活性。一般认为它由大小2个亚基组成。大亚基为贯穿膜全层的脂蛋白,为催化部分;小亚基为细胞膜外侧的糖蛋白,其作用机制尚不清楚,但如果将大小亚基分开,酶活性即丧失。在大亚基的细胞质端有与 Na+ 和 ATP结合的位点,外端有乌本苷 (ouabain)的结合点,可以反复发生磷酸化和去磷酸化。乌本苷为 Na+/K+ - ATPase酶的抑制剂。
Na+/K+ -ATPase酶是一种离子泵,能分解消耗1个分子 ATP能量将3个 Na+ 从细胞内低浓度侧转运到细胞外高浓度侧,同时转运2个 K+ 泵从细胞外低浓度侧转运到细胞内高浓度侧。具体过程为:①Na+ 和 ATP与酶结合;②酶的活性激活后,分解ATP,产生的高能磷酸根使酶发生磷酸化;③酶构象改变,将 Na+ 结合部位暴露到膜外侧,此时酶对 Na+ 的亲和力变低;④Na+ 被释放到细胞外的同时,酶对 K+ 的亲和力增高,K+ 结合到酶上;⑤K+ 的结合促使酶发生去磷酸化;⑥酶去磷酸化后构象复原, K+ 结合部位转向膜的内侧,这时的酶与 Na+ 的亲和力高,与 K+ 的亲和力变低,因而在膜内侧释放 K+ ;⑦Na+/K+ -ATPase恢复至初始状态。如此反复进行的构象变化每秒钟可完成1000多次 (图4)。细胞内约有1/3以上的能量 (ATP)是被 Na+/K+ -ATPase活动消耗的。Na+/K+ -ATPase在维持膜电位、调节渗透压、控制细胞容积和驱动糖与氨基酸的主动运输等方面都起着重要的作用。另外,胃中还有一种质子泵为 H+/K-+ ATP酶,这些质子泵负责创造胃的酸性环境,并且可以引起酸回流。 治疗胃酸过高的药物如奥美拉唑 (omeprazole)等质子泵抑制剂可用于溃疡或酸反流患者,以帮助降低其肠道酸度。
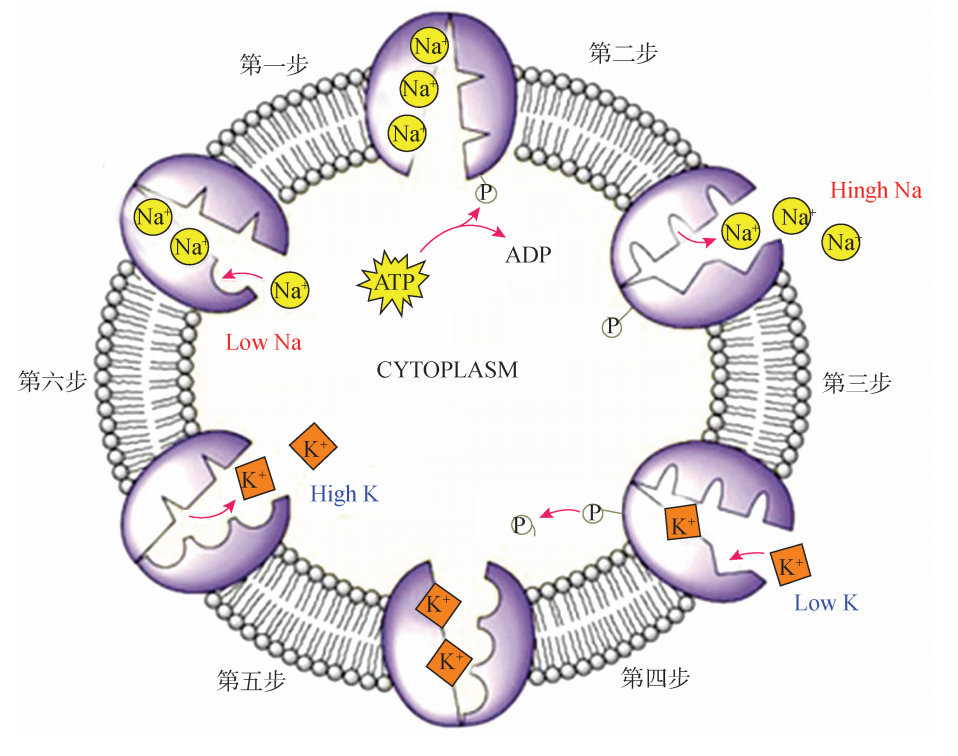
图4 Na+/K+ -ATPase酶转运 Na+ 和 K+ 的过程
(2)次级主动运输:次级主动运输又称为继发性主动转运、联合转运、协同运输. 某种物质能够逆浓度差进行跨膜运输,但是其能量不是来自 ATP分解,而是由联合转 运的其他离子的高势能提供.沿着其电化学梯度向下移动的离子被称为驱动离子.与 初级主动运输不同,在次级主动运输中,执行主动运输所需的自由能由驱动离子的浓 度梯度提供.Na+ 在许多细胞的质膜中充当许多 (但不是全部)次级主动运输蛋白的 驱动离子,其 Na+ 电化学梯度是由 Na+/K+ -ATPase来保持的..
例如,小肠上皮细胞和肾细胞能利用 Na+ 跨膜梯度驱动转运,特异性地吸收氨基 酸和葡萄糖.这种运输过程中伴有 Na+ 进入细胞 (图5).具体地讲,这种过程是 由膜上的 Na+/K+ -ATPase和特异性的载体蛋白共同协作完成的.载体蛋白上具有2 个结合位点,可分别与 Na+ 和葡萄糖 (或氨基酸)结合.当 Na+ 顺浓度梯度进入细胞 时,葡萄糖或氨基酸就利用 Na+ 的势能驱动,随着载体蛋白构象变化,与 Na+ 相伴逆 浓度梯度进入细胞,这是一种共运输.Na+ 浓度梯度差越大,葡萄糖或氨基酸等物质 进入细胞的速度就越快,Na+/K+ -ATPase则靠分解 ATP提供能量,再把 Na+ 泵出 细胞外,维持 Na+ 的浓度梯度。
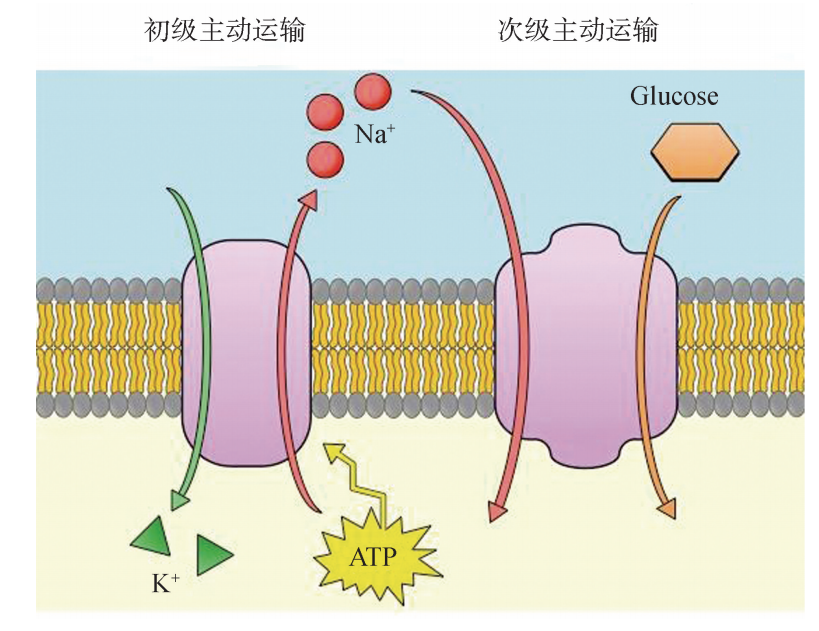
图5 Na+ 浓度梯度驱动下的次级主动运输
主动运输的载 体 蛋 白 同 时 转 运 的 两 种 物 质 若 沿 同 一 方 向 转 运,称 为 同 向 运 输 (symport);如果这两种物质朝相反的方向转运,则称为反向运输 (antiport).