
第6章 森林群落演替
Chapter 6 Succession of Forest Communities
【本章提要】森林群落发生的过程包括迁移、定居、竞争和反应4个阶段。森林群落演替可根据不同原则划分为不同演替类型。演替顶极学说主要有单元顶极论与多元顶极论。本章介绍了森林动态模型及林窗动态模拟方法,叙述了恢复生态学基本原理和生态恢复关键技术。
6.1森林群落发生、发育的一般过程
6.1 Normal process of forest community development
6.1.1森林群落发生的过程
森林群落的发生一般都具有迁移→定居→竞争→反应这几个过程。
(1)迁移:从繁殖体开始传播到新定居的地方为止的过程。
(2)定居:繁殖体迁移到新的地点后,进入定居过程。定居包括发芽、生长、繁殖三个环节。各环节能否顺利通过,取决于种的生物学、生态特征和定居地的生境。
(3)竞争:在一定的地段内,随着个体的增长、繁殖,或不同种的同时进入,必然导致营养空间和水、养分等和竞争,结果是“最适者生”。竞争的结果,使森林群落随年龄的增加单位面积上林木株数不断减少,所谓森林群落“自然稀疏”。
(4)反应:通过定居过程,群落内生物与非生物环境间的能量转换和物质循环不断进行,原来的生境条件逐渐发生相应的变化,这就是“反应”。
在自然界中,上述过程经常交织在一起,不易截然分开。一般来说,迁移和定居是顺序进行的,而竞争与反应则基本与定居同时发生,只不过初期在程度上不是那么激烈或明显。
讲解论文:
[1]杨 宁, 陈 璟, 杨满元, 等. 贵州雷公山秃杉林不同林冠环境下箭竹分株种群结构特征[J]. 西北植物学报, 2013, 33(11): 2326-2331.
[2]杨 宁, 付美云, 杨满元, 等. 衡阳紫色土丘陵坡地不同土地利用模式下土壤种子库特征[J]. 西北植物学报, 2014, 34(11): 2324-2330.
[3]杨 宁, 邹冬生, 杨满元, 等. 贵州雷公山秃杉的种群结构和空间分布格局[J].西北植物学报, 2011, 31(10): 2100-2105.
[4]杨 宁, 邹冬生, 李建国, 等. 衡阳盆地紫色土丘陵坡地主要植物群落自然恢复演替进程中种群生态位动态[J]. 水土保持通报, 2010, 30(4): 87-93.
[5]杨 宁, 邹冬生, 李建国. 衡阳盆地紫色土丘陵坡地自然恢复灌丛阶段主要种群空间分布格局[J]. 生态环境学报, 2009, 18(3): 996-1001.
6.1.2森林群落发育的时期
一般划分为3个时期:发育初期→发育盛期→发育末期。
6.2森林群落演替的主要类型
6.2 Main patterns of forest community succession
6.2.1按初始生境水分条件划分
(1)旱生演替:地衣植物群落阶段→苔藓植物群落阶段→草本植物群落阶段→木本植物群落阶段。
(2)水生演替:沉水植物群落阶段→漂浮植物群落阶段→苇塘阶段→苔草草甸阶段→疏林阶段→中生森林阶段。
(3)中生演替:裸露矿质土阶段→草本植物阶段→木本植物阶段。
6.2.2按演替起始裸地性质划分
(1)原生演替:开始于原生裸地上的植物群落演替。
(2)次生演替:开始于次生裸地上的植物群落演替。次生演替一般具有以下特征:a. 次生演替发生的动力来源于外部的干扰,人为或自然干扰均能消除原有植被,从而发生演替。次生演替起点的生境因保留原群落繁殖体也能向裸地提供种源,所以次生演替能较快地重新被植被覆盖,演替速度较快,当然这决定于干扰的类型、频度和强度等;b. 干扰因素一停止,演替发展途径有3种(促进途径,忍耐途径和抑制途径)。
6.2.3按演替延续的时间划分:(1)世纪演替;(2)长期演替;(3)快速演替.
6.2.4按控制演替的主导因素划分:(1)内因性演替;(2)外因性演替.
6.2.5按演替方向划分:(1)进展演替(Progressive succession);(2)逆行演替(Regressive succession或Retrogressive succession);(3)循环演替。
6.3演替顶极学说
6.3 Succession climaxes
演替顶极学说最早是由英美学派提出的,近几十年来,得到不断修正、补充和发展。有关演替顶极理论主要有3种:单元顶极论、多元顶极论和顶极—格局假说。
6.4森林演替实例
6.4 Case studies of forest community succession
6.4.1亚高山暗针叶林区泥石流迹地植被原生演替:川滇柳、冬瓜杨幼苗群落→川滇柳、冬瓜杨幼树群落→川滇柳、冬瓜杨林→冬瓜杨、峨眉冷杉林→峨眉冷杉中龄林→峨眉冷杉成熟林→峨眉冷杉过熟林。
6.4.2阔叶红松林的演替
![]()

6.4.3采伐迹地的演替
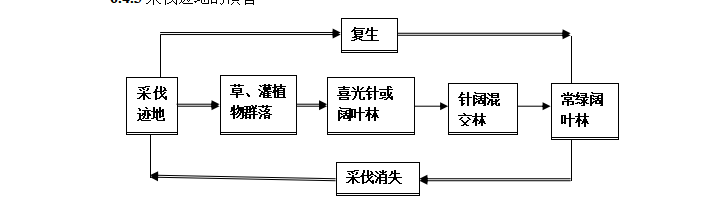
![]()
6.5森林动态模型
6.5 Dynamic models of forests
6.5.1概述
经典的森林演替研究主要是以追溯法和用空间代替时间的方法为主。这些方法的据点是研究周期长,而且,难于对森林未来的动态变化作出定量预测。随着20世纪60年代计算机技术的发展,生态学家能够用经典的数学问题表达自己的思想,使森林动态模拟得到飞速发展,对于揭示森林动态发展机制,提供了一种重要的工具。森林动态模拟通常可分为林分总体模拟;树种更替模拟;直径转移模拟;空间竞争模拟和林窗动态模拟5大类。
6.5.2林窗动态模拟(Gap-phase Dynamic Modeling)
(1)林木最优生长的模拟
在KOPIDE模型中,用Richards方程表示树高—胸径关系:
![]()
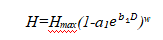
式中:a1, b1, w——待定参数。
胸径与叶量的关系为:
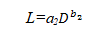
式中:a2, b2—待定参数。
(2)环境因子对林木生长的限制作用
a. 光与林木生长的关系
阳光与林冠层的投射服从于Beer-Lambert定律:
Qh=Q0e-kL(h)
式中:Qh—树高h处的透光量(0~1);Q0—树冠上方的入射量;L(h)—在树高h上方的累积叶面积指数;
k与树种组成及地理纬度有关。在KOPIDE模型中,k=0.25。根据光合产量与光强的一般关系,林木生长与透光量的关系为:
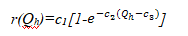
式中:r(Qh)—光照限制因子(0~1);c1—尺度常数;c2—曲率常数;c3—林木的光补偿点。
b. 土壤湿度及肥力与林木生长的关系
KOPIDE模型用抛物线方程表示湿度限制因子:
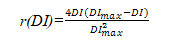
式中:r(DI)—土壤湿度限制因子(0~1);DI—干燥度(年蒸发量与年降水量之比);DImax—树木分布区的最大干燥度。
c. 气温与林木生长的关系
气温限制因子是根据年积温确定的,林木的生长对气温的反映可用抛物线函数表示:
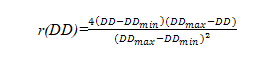
式中:r(DD)—气温限制因子;DD—大于10 ℃或5 ℃的年积温;DDmin,DDmax—树木分布范围内最小和最大的年积温。
(3)林木更新的模拟
在KOPIDE模型中,采用的是林冠叶面积指数,最大可能的林木更新株数与林冠层的叶面积指数的关系可用指数函数表示:
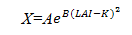
式中:X—林木更新株数;LAI—林冠层叶面积指数;B—与最大有效叶面积指数有关的待测参数;K—最佳叶面积指数。
(4)林木死亡的模拟
林窗模型把树木的死亡视为随机事件,根据树木死亡的原因分为自然死亡和不良环境引起的死亡。自然死亡由两个假设条件来计算:1)只有1%的树木能够生长到最大树龄;2)每年死亡的概率相同。
假设林分内共有N株林木,T年后,只余下n株,树木死亡率为pm,当T=n时有:
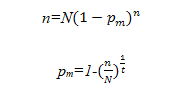
当n/N=0.01,T=Amax时有:
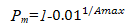
近似计算得:
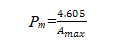
不良环境对林木造成影响直到死亡可从林木生长量反映出来。在林木死亡前,林木胸径的连年生长量逐渐减小。当生长量达到临界生长量后,在一定年限生长条件不能得到改善,必然死亡。Pacala等把林木的死亡直接与生长量联系起来:
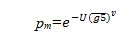
式中: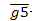 —最近5 年林木生长的算术平均值;U, V—待定参数.
—最近5 年林木生长的算术平均值;U, V—待定参数.
6.6恢复生态学原理
6.6 Principles of restoration ecology
6.6.1恢复生态学概念
6.6.2恢复生态学的理论基础
6.6.3生态恢复的过程
生态恢复的一般过程是:本底调查→区域自然、社会经济条件(水、土、气候、可利用的条件等)综合分析→恢复目标的制定→恢复规划→恢复技术体系组配→生态恢复实施→生态管理→生态系统的综合利用→自然—社会—经济复合系统的形成。
6.6.4生态恢复的关键技术
参考以下文献文献讲解:
[1]杨 宁, 邹冬生, 李建国. 衡阳盆地紫色土丘陵坡地植被恢复模式建设[J]. 草业科学, 2010, 27(10): 10-16.
[2]杨宁, 彭晚霞, 邹冬生, 等. 贵州喀斯特土石山区水土保持生态经济型植被恢复模式[J]. 中国人口.资源与环境, 2011, 21(S1): 474-477.
6.7天然林保护工程
6.7 Natural forests conservation project
6.8退耕还林工程
6.8 Grain-to-forest
复 习 思 考 题
(1)森林群落的发生进程包括哪几个阶段?各自特点如何?
(2)旱生演替系列包括哪4个阶段?
(3)水生演替系列包括哪6个阶段?
(4)举例说明次生演替的一般特征。
(5)森林动态模拟通常分哪几种类型?
(6)生态恢复的一般过程有哪些步骤?
本 章 推 荐 阅 读 书 目
[1]蒋有绪, 郭泉水, 马 娟, 等. 中国森林群落分类及其群落学特征[M]. 北京: 科学出版社, 1998.
[2]张佩昌, 周晓峰, 王凤友. 天然林保护工程概论[M]. 北京: 中国林业出版社, 1999.
[3]李育才. 中国的天然林资源保护工程[M]. 北京: 中国林业出版社, 2004.
[4]曲仲湘, 吴玉树, 王焕校, 等. 植物生态学(第2版)[M]. 北京: 高等教育出版社, 1983.

