
第二节 群落的种间关系
生物群落中物种之间的关系是十分多样而复杂的。各种生物互为环境中的生物因子,它们之间互相联系,互相影响,形成各种种间关系。
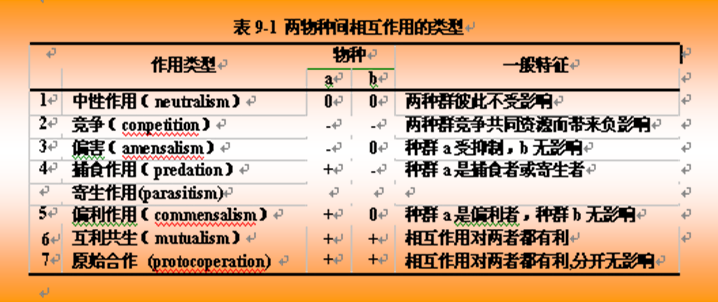
生物群落中物种形成种间关系(interspecific relationship)。种间关系主要表现在营养关系(食物关系)上,具体体现在竞争与中立;捕食与寄生;共生互利;偏利和偏害;刺激与抑制。
下表两物种之间相互作用的基本类型。表中“+”表示对存活或其他种群特征有益;“-”表示对种群增长或其他特征有抑制;而“0”表示无关紧要的、没有意义的相互影响。
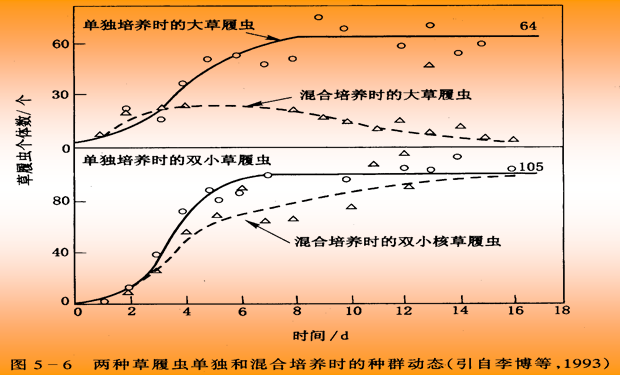
竞争是指两个或多个种群当所需的环境资源或某种必需的环境条件受到限制时所发生的相互关系。能充分利用环境资源和能量的物种较好的生长,其它物种得到的资源和能量不足,生长和生存都受到限制。
竞争的结果,生态位相同的两个种群不能永久共存, 这就是高斯原理( Gause’s principle),又称竞争排斥原理(competitive exclusion principle)。
竞争与生态位参考学习资料
生态位(niche)是指一种生物在生物群落 (或生态系统)中的功能或作用。也是指某一物种的个体与环境(包括生物环境和非生物环境)之间特定关系的总和。它包括: 空间、营养、超体积生态位 。
生态位的概念最早由Grinell (1917)提出,用来表示对栖息地进行划分的空间单位,即相当于空间生态位(spatial niche),有的学者译为“小生境”但这易与生境(habitant)——生物的栖所相混淆。
1927年,Elton将生态位定义为物种在生物群落中的地位和“角色”(role),主要是指营养生态位(trophic niche)而言的。
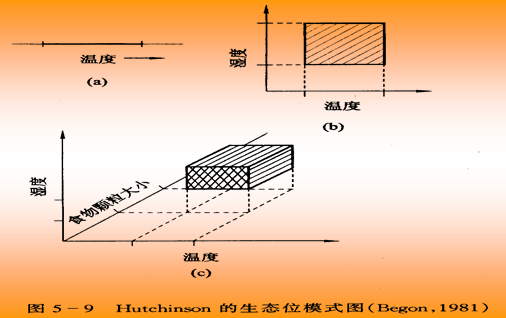
超体积生态位
Hutchnson (1958)指出,某种生物的生态位是由所有生物和非生物环境条件组成的n维空间中该生物得以继续生存的范围即超体积生态位如某种生物在一定的温度范围内生存、生长和繁殖,则这个温度范围就是该种生物的一维生态位。加上其他条件如水深、食物颗粒大小,这样就形成二维和三维生态位(图9-1)。如将生物要求的几个条件都考虑进去,就构成一个抽象的n维空间,这个超体积 当然无法直接表示,但可以列成表格或用多角图形反映出来。
此外,Hutchnson (1965) 还进一步区分为基础生态位和实际生态位,前者指某一物种所栖息的理论上的最大空间,后者则是在竞争和捕食等压力下某一物种实际占有的空间。
竞争种类越多,使某物种占有的实际生态位可能越来越小。Hutchnson提出的生态位内容相当完整,但要了解生物所需的全部条件是件难事,因此,在操作上,不妨以Elton的生态位即生物的功能地位或“职业”的观点为基础,选择生物所需条件中的几个可量因素去研究生态位的问题。如决定一个水生物种正常繁殖的各种环境变量包括:温度、溶氧、盐度、pH、水流以及食物的种类、大小和组成等因素,物种对这些因素的适宜范围综合构成物种能够栖息生存的多维(变量)空间,即生态位。
如果两个物种在一个稳定的群落中具有相同的基础生态位,即生态位完全重叠,那么两个物种不能共存,竞争优势种会把另一物种完全排除掉或迫使它改变生态位;如一个物种的基础生态位可能被完全包围在另一个物种的基础生态位之内,在这种情况下,竞争结果取决于两者的竞争能力。若生态位被包在里面的物种处于竞争劣势,它会消失;若里面的有竞争优势,它就会把外包物种从发生竞争的生态位空间中排挤出去,从而实现两个物种共存。
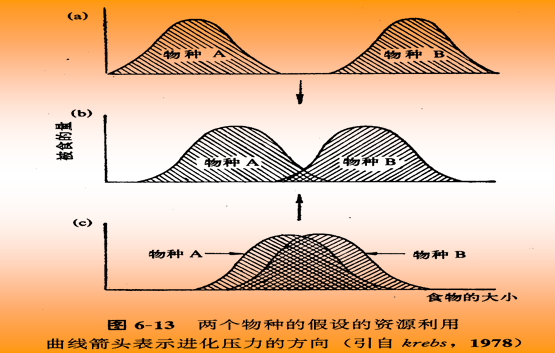
在一个稳定的群落中,所有物种的实际生态位各有所不同。例如,两个物种的基础生态位若发生部分重叠,即有一部分生态位是被两个物种共同占有,其余部分各自分别占据,则会产生局部竞争,这时具有竞争优势的物种会占有重叠部分的空间而成为优势种;较弱的竞争者数量减少,但种群仍能存在。因此,两个物种实现共存。
基础生态位与实际生态位
(1)基础生态位:
指生物群落中,某一物种所栖息的理论上最大空间,即没有种间竞争的种的生态位。
(2)实际生态位:
指当有竞争者时,必定使该物种只占据基础生态位的一部分,这一部分实际占有的生态空间称为实际生态位。 竞争种类越多,某物种占有的实际生态位越小。
实际生态位是生物之间的相互影响和物种自己对某些环境条件所具有选择性的共同结果
该概念对正确认识物种在自然选择进化过程中的作用,以及运用生态位理论指导人工群落建立中种群的配置等方面具有十分重要的意义。
生态位分类系统
(1)每一种生态因子对应着一种或一维特定的生态位,如光、温度、食物等生态位;
(2)按照生态元的类别,有基因、细胞、个体、物种、生态系统、城市、生物圈、地球生态位;
(3)根据竞争与否,分基础生态位和实际生态位,前者包括后者;
(4)生态位如果由生态元自身生产而形成的称为自产生态位,如鸟窝、人类建造的房、生态的粮食等;
(5)由其他生态元产生或自然存在、发生的生态位称非自产生态位
空间生态位:
表示划分环境空间单位和一个物种在环境中的地位。一个物种所占有的微环境。
营养生态位:
表示物种在生物群落中的地位与功能作用,强调的是物种与物种之间的营养关系。
这是一张植物根系资源分化,三种不同植物根在同一土壤区域同存。荨麻、印度锦葵、狗尾草.
![]()
(1) 一个稳定的群落中占据了相同生态位的两个物种,其中一个终究要灭亡;
(2)一个稳定的生物群落中,由于各种群在群落中具有各自的生态位,种群间能避免直接的竞争,从而保证了群落的稳定。
(3)群落是一个相互起作用,生态位分化的种群系统。这些种群在它们对群落的时间、空间和资源利用方面,以及相互作用的可能类型方面,都趋于互相补充而不是直接竞争。大家配合共同生活,更有效地利用环境资源,从而保证了群落在一个较长时间有较高的生长力,具有更大的稳定性。
(4)竞争可以导致多样性而不是灭绝,竞争在塑造生物群落的物种构成中发挥着主要作用。竞争排斥在自然开放系统中,很可能是例外而不是规律,因为,物种常常能够转换它们的功能生态位去避免竞争的有害效应。
![]()
生物多样性的分布格局和维持机制关键是物种的共存机制。
长期以来,生态位分化的思想在这一研究领域占据着主导地位。然而这一理论在解释热带雨林很高的物种多样性时遇到了困难。
由于生态位理论的不足,与实际情况似乎存在矛盾:
浮游生物悖论:环境条件非常均一的水体内共存着至少数十种浮游植物.
热带雨林:太高的物种多样性,无法确定每种植物的生态位。
由于生态位理论不能给这些群落学模式提供令人满意的解释。近年来中性理论应运而生。以美国生态学家Hubbell为代表的学者,认为:
(1)生态学上相同的物种可以实现共存,物种多度的变化是随机的,而非确定性的;
(2)共存的物种数量取决于物种分化和随机灭绝之间的平衡;
(3)群落内物种的相对多度随时间表现为随机振荡的波动。
捕食
1.捕食者与猎物的关系
除了竞争食物和空间等资源外,物种之间还有直接的捕食现象。通俗地说,捕食就是一种生物吃另一种生物,可以是杀死猎物,也可能不杀死。广义的捕食可以包括四种类型:
典型的捕食是指食肉动物吃食草动物或其他食肉动物。侠义的捕食者与 猎物就是这种典型捕食的两个物种。
寄生现象也是一种捕食现象。
同类相食(cannibalism)是一种持殊的捕食现象。即捕食者与猎物属于同一种类,这种现象在水生生物中是常见的
牧食,即食草动物吃绿色植物,这种情况下植物往往未被杀死而受损害。
他感作用(allelopathy)
他感作用(allelopathy),是指植物植株向环境中释放某些化学物质,影响周围其它植株生理生化代谢及生长过程的现象。
具有他感作用的物质称作他感化合物。如该物质直接来源于植物的分泌或分解出来的,其产生的他感作用称为真他感;如他感化合物是通过微生物降解来的,间接来源于植物则称为次生他感化合物,其作用称为功能性他感。
他感作用是自然界存在的一种普遍现象,它既存在于不同杂草种群之间,如小飞蓬产生 C10 -聚乙炔甲酯抑制豚草种子发芽,也存在于杂草与作物之间,如野燕麦的根系分泌出莨菪亭及香草酸等抑制小麦的生长发育,小麦的根系分泌物抑制白茅的生长。还存在于杂草同种不同个体或作物与作物之间,如小飞蓬根腐烂产生的他感作用抑制其幼苗的生长,腐烂的小麦残体抑制玉米的生长。老桃园残留桃树皮中扁桃苷的降解产物氰化物对新种植的桃树有毒害作用。同种植物不同个体间的他感作用,常称作“自毒作用”。
此外,他感作用还指促进植物生长的一面,如麦仙翁产生的麦仙翁素可促进小麦的生长,也是他感作用的表现。
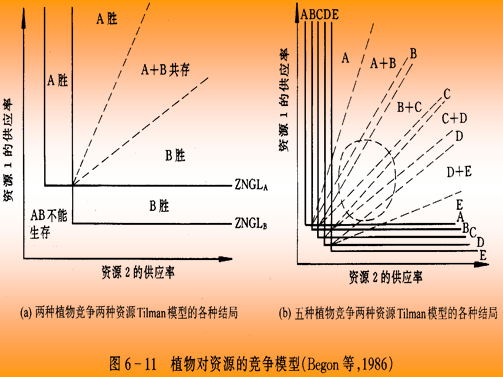
种群竞争模型
![]()
假设两个物种,当它们单独生长时增长曲线符合逻辑斯谛模型。
dN1 /dt = r1N1 (1-N1/K1)
dN2 /dt = r2N2 (1-N2/K2)
如果将这两个种放在一起时,它们之间会出现(资源、食物、空间的)竞争。
假设两个参数:α、 β
α表示在物种1的环境中,每存在一个物种2的个体,对于物种1产生的效应。
β表示在物种2的环境中,每存在一个物种1的个体,对于物种2产生的效应。
则有逻辑斯谛方程:
dN1 /dt = r1N1 (1-N1/K1-αN2/K1)
dN2 /dt = r2N2 (1-N2/K2-βN1/K2)
各参数的含义:
其中:
N1、N2:分别为两个物种的种群数量
K1、K2:分别为两个物种的环境容纳量
r1、r2 : 分别为两个物种的种群增长率
α、β:物种2对物种1的竞争系数,物种1对物种2的竞争系数。
![]()
从理论上看,物种1和物种2的竞争结果是由两个物种的竞争系数α、β和环境容量K1、K2共同决定的。
这里存在四中不同的情况:
1.α>K1/K2和β<K2/K1
1/K为种内竞争指标,
α/K1为种群2对1的种间竞争强度
β/K2为种群1对2的种间竞争强度
α>K1/K2,即:α/K1>1/K2
它表示:物种2中的每个个体对物种1的影响>物种2中每个个体对自身的影响
β<K2/K1,即:β/K2>1/K1
它表示:物种1中每个个体对物种2的影响<物种1中每个个体对自身的影响
这就使得物种1处于劣势,物种1被排斥,物种2 取胜
2.α<K1/K2和β>K2/K1
这个情况就跟第一种情况正好相反:
物种2中的每个个体对物种1的影响<物种2中每个个体对自身的影响。
物种1中每个个体对物种2的影响物种>1中每个个体对自身的影响
使得物种2处于劣势,物种2被排斥,物种1取胜
3.α>K1/K2或β>K2/K1
物种2中的每个个体对物种1的影响>物种2中每个个体对自身的影响
物种1中每个个体对物种2的影响>物种1中每个个体对自身的影响
此时:种间竞争>种内竞争。平衡点不稳定,两个物种都有可能获胜。
4.α<K1/K2和β<K2/K1
物种2中的每个个体对物种1的影响<物种2中每个个体对自身的影响
物种1中每个个体对物种2的影响<物种1中每个个体对自身的影响
此时:种间竞争<种内竞争。平衡点稳定,两个物种共存。
![]()
通过分析一年生植物的竞争关系,可以提出一年生植物的竞争模型。
根据播种时及收获时种子数的比例进行分析:
大麦和燕麦竞争试验,实验结果如下:
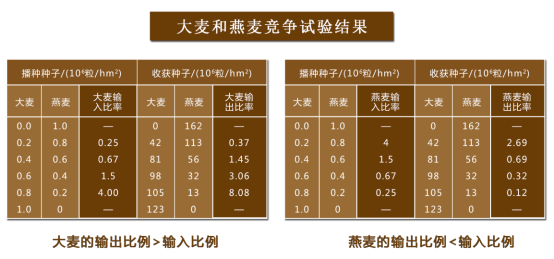
可得结论:
(1) K1>K2/β, K1/α> K2, 物种2被排斥,物种1取胜;1/K1<β/K2, 1/K2 >α/K1
(2)K1/α<K2, K1<K2/β 物种1被排斥,物种2 取胜;1/K1>β/K2, α/K1>1/K2
(3)K1/α<K2或 K1>K2/β,物种1、2都可能获胜; β/K2 > 1/K1, α/K1>1/K2
(4) K1/α>K2, K1<K2/β两种共存,达到某种平衡。 1/K2>α/K1, 1/K1>β/K2
竞争排斥原理的反对派认为,许多亲缘很近的种类往往在同一栖息地中生活在一起。
一种认为,自然界中很少见到竞争,即物种很少为有限资源而竞争。因此,在自然群落中没有必要去找竞争排斥的证据。
另一种观点认为,自然界中。竞争十分普遍,它是群落中支配物种进化发展的主要因素。那些在同样的环境中能够共存的近缘种,其生态位有明显区别的,竞争由此而减少.

