第七节 物种形成与种群生态对策
一、物种形成
选择的结果形成了生物的适应性。不过适应性并未创造出“最好”或“最优”的表现型,这是因为自然选择只作用于现有的表现型,同时任何适应性都是相对的:
种群经常出现基因突变(mutation)
环境不断变化,原来适应于某一特定环境的表现型变成了不适应的。
在有限的能量和资源条件下,有机体必须把所获取的能量和资源分配给一系列彼此矛盾的目的,包括生长、生殖、维持、防卫、修复、贮存等,因此常采取调和的对策。
选择进化的关键阶段是新物种的形成,目前广为学者们所接受的是地理物种形成(分三步):
地理隔离--通常由于地理屏障引起,将两个种群彼此隔开,阻碍了种群间个体的基因交流
独立进化--两个地理与生殖上隔离的种群各自独立发展变化,适应各自独立的特殊环境
生殖隔离机制建立—若以后隔离屏障又消失,两个种群的个体可再次相遇和接触,然而因久远隔离已使其无法再基因融合--新种从而形成
物种形成方式
1.同域性:生活在同一区域内的物种, 由于资源的限制和种群内部的激烈竞争,导致生态位出现分化。占据不同生态位的群体出现基因交流的障碍,通过生殖隔离而形成新的物种。
2.邻域性:有些物种的分布区很广但扩散能力较差,在其分布区的边缘地带的一些种群,由于栖息地环境的差别而形成基因交流的阻碍,逐渐建立起自己独特的基因库,并形成生殖隔离,最终形成了新的物种。
3.异域性:一个物种的多个种群生活在不同的空间范围内,由于地理隔绝使这些种群之间的基因交流出现障碍,导致特定的种群积累着不同的遗传变异并逐渐形成各自特有的基因库,最终与原种群产生生殖隔离,形成新的物种。
地理变异 (geographic variation)--广布种的形态、生理、行为和生态特征往往在不同地区有显著的差异,称~~,它反映出物种种群对环境选择压力空间变化的适应反应
渐变群 (cline) --选择压力在地理空间上的连续变化,导致基因频率或表现型的渐变,形成一变异梯度,称~~
地理亚种 (subspecies)--选择压力在地理空间上不连续变化下,形成的变异种群
二、种群的数量变动
任何一种生物种群,都有不同程度的数量和生物量的变动。环境的变化越剧烈,种群数量的变化幅度也越大。当生活条件优良时,有机体不仅个体数量增加,而且生长加快 ,因而种群生物量也急剧增大;当生活条件恶化时,数量和生物量则降低,也就是说种群的再生产能力缩减到与具体条件相适应的水平。但是种群数量变化不是环境因素单方面作用的结果。各种生物都形成按照环境变化而调节的内源性反馈机制,数量变动是环境因素和种群适应性相互作用的结果。
(一)种群数量的日变化
(二)、种群数量的季节消长
(三)种群数量的逐年变化
(四)种群数量的非周期性波动
三、种群的生态对策:r -对策者和K-对策者
任何一种生物的种群,都有不同程度的数量和生物量的变动。各种生物都能按照环境的变化而调节种群的数量,数量变动是环境因素和种群适应性相互作用的结果。
任何生物种群都具有相对稳定性。表现在种群的变动具有一定的上限和下限,还可能有一个长时间的平均水平(时滞)。当种群密度离开平均水平上升或下降时,就有一种使种群密度重返原来水平的倾向。并且种群数量上升或下降越剧烈,这种相反的倾向也越显著(即反馈调节)。
在复杂的自然环境条件下,各种生物都具有其独特的出生率,寿命,形体 大小,存活率等生态特征。这些相互联系的生态特征,是物种在长期的进化过程中,与环境相互作用并通过自然选择而形成的。在生物进化过程中,按种群的动态类型形成了两类互不相同的适应。
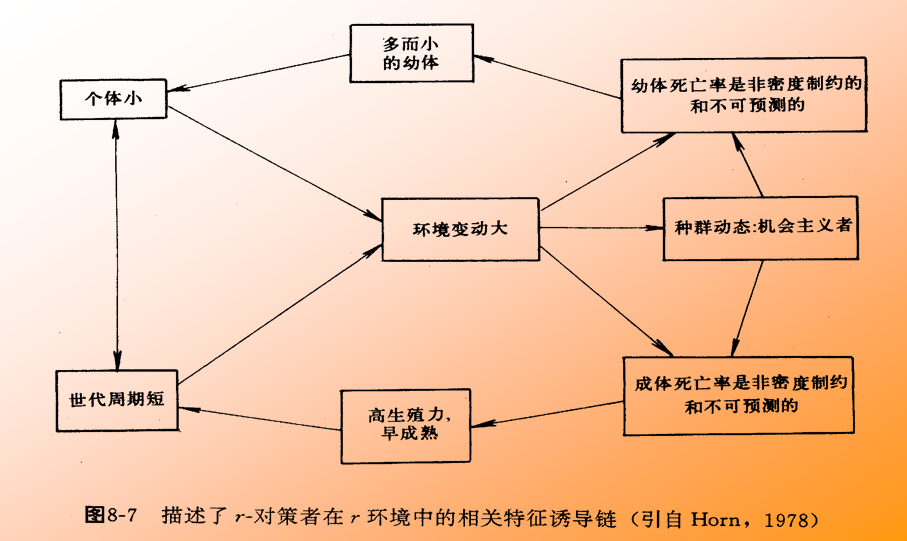
一类称为r-选择(r-selected):这类生物它们的种群密度很不稳定,很少达到环境所能负载的最高负荷量——K值。一般保持在 “ S ”型生长曲线的上升段。这类生物通常出生率高,寿命短,个体小,常常缺乏保护后代的机制, 子代死亡率高,但具有较大的扩散能力。它们适应于不稳定的生活环境。
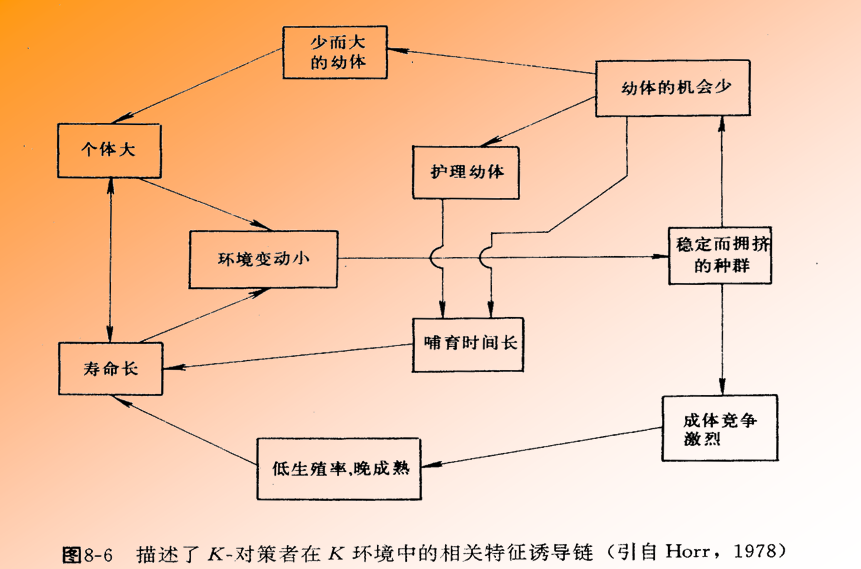
另一类称为K-选择(K-selected):它们的种群密度比较稳定,经常处于K值周围,这类生物通常出生率低,寿命长,个体大,具有较完善的保持后代 的机制,子代死亡率低,但一般扩散能力也较低。它们适应于稳定的生活环境.
从进化论观点来看,生物适应于不同栖息环境并朝着不同方向进化的“对策”称为生态对策(bionomic strategy),属于r-选择的生物可称为r-对策者(r-strategists);属于K-选择的生物可称为K-对策者(K-strategists)。
r-对策者和K-对策者是两个进化方向不同的类型,在这中间还有一系列过渡的类型,有的更接近于r-对策者,有的更接近于K-对策者,也就是说从典型的r-对策者到 典型的K-对策者之间有一个连续的谱系。
K-对策者的栖息环境是稳定的,在这种条件下增加种间斗争和竞争能力使种群密度保持在饱和密度附近是有利的,因此被选择的物种常常是个体大者。大个体由于增加世代周期而降低出生率,而出生率的降低必定要相应的增加存活率。因此这类动物防御和保护幼代的能力一般比较强。由于有亲代的抚育,出生率又低,也更容易达到长寿命和大的个体。K-对策者种群通常由许多不同的年龄世代组成,某一世代数量的变动可以从其它世代中得到平衡,但是由于r值较小,当种群遭受过度的死亡或激烈的动乱后,返回平衡水平的能力是比较低的,因此容易灭绝,对于这一类动物的资源应更加注意保护。
r-对策者的栖息环境是不稳定的﹑多变的,因而种群密度也难免处于经常激烈变动状态。个体小﹑寿命短有利于高的r 值,能使它们在有利条件下 迅速占领生境。因为它们占有的生境常是生态真空的,因而竞争能力不发达,对捕食者的防御能力比较弱,死亡率很高。但它们的高出生率可以弥补死亡的消耗。当种群密度过高和把资源耗尽时,这类动物具有较强的扩散能力,迁移恶化的生境到别处建立新的种群;有的种群则以休眠形式潜伏下来。