三、染色质组装形成染色体

(一)核小体—染色质的一级结构(基本结构单位)
核小体(nucleosome):是染色体的基本结构单位,为由200bp左右的DNA分子及一个组蛋白八聚体构成的圆盘状颗粒。
组装:核小体组蛋白H2A 、 H2B、H3和H4各两分子组成八聚体;146bp的DNA分子盘绕组蛋白八聚体1.75圈,形成核小体;两个相邻核小体之间以连接DNA相连,典型长度为60bp;组蛋白H1结合于连接DNA,位于核小体核心DNA双链的进出端,起稳定核小体的作用。核小体串珠的形成使DNA分子压缩了约7倍。
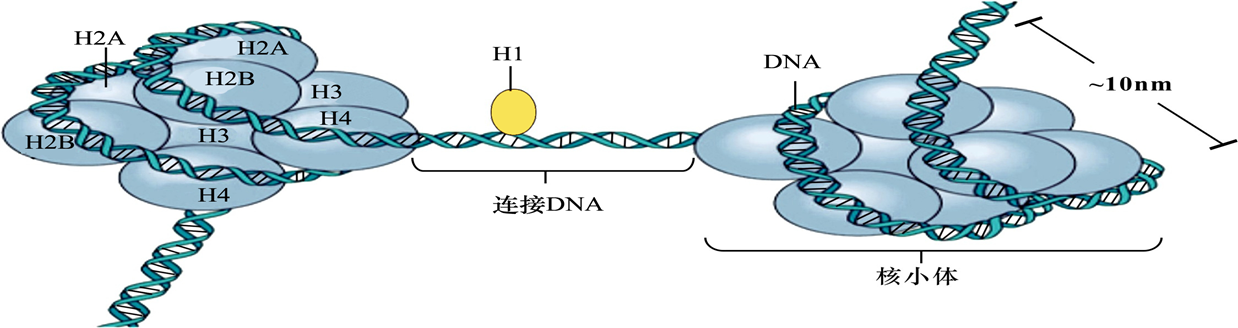
图8-15 核小体结构示意图
(二)螺线管———染色质的二级结构
概念:螺线管(solenoid)是在组蛋白H1协助下,由核小体串珠结构盘旋而成的中空结构。
组装:螺线管每圈6个核小体,螺距11nm,外径30nm,内径10nm。
组蛋白H1:由一个球形中心及两个氨基酸臂组成,球形中心可与核小体上的特异性位点结合,两个臂则与相邻核小体组蛋白核心上相应的位点结合,以协助核小体包装成有规律的重复排列结构。螺线管的形成使核小体串珠结构压缩了约6倍。
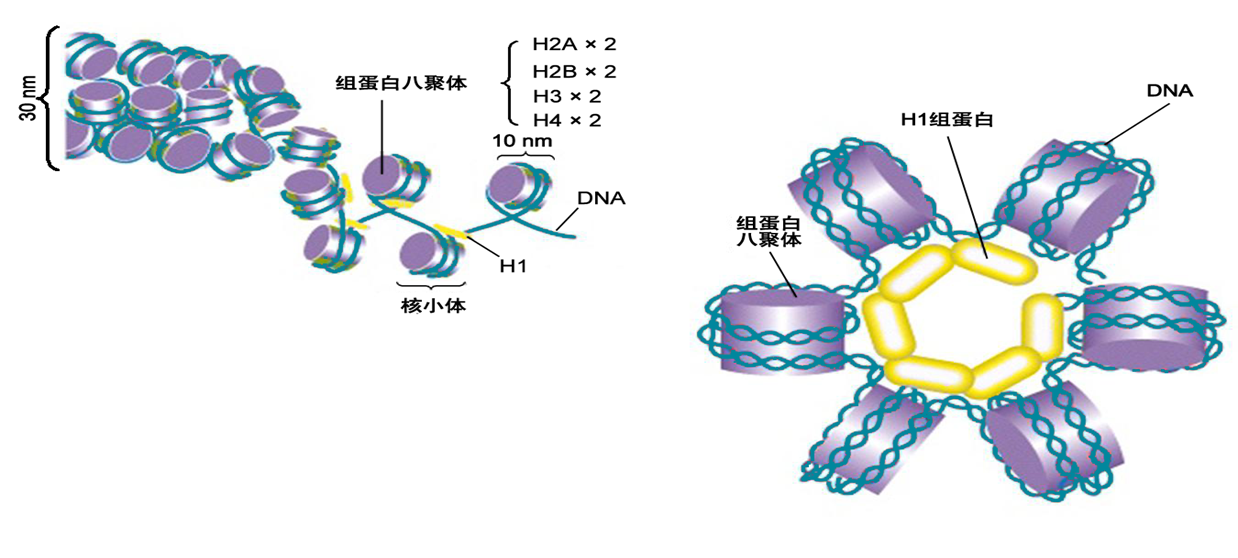
图8-16 螺线管结构示意图
(三)螺线管的进一步包装
关于螺线管如何进一步包装成染色体,目前有两种模型:多级螺旋化模型和染色体骨架-放射环模型。
1.多级螺旋化模型(multiple coiling model)
一级结构——核小体
二级结构——螺线管
三级结构——超螺线管(supersolenoid):由螺线管进一步螺旋化形成的圆筒状结构
四级结构——染色单体(chromatid):超螺线管进一步螺旋折叠形成。
DNA→核小体→螺线管→超螺线管→染色单体。分别压缩7倍、6倍、40倍、5倍。
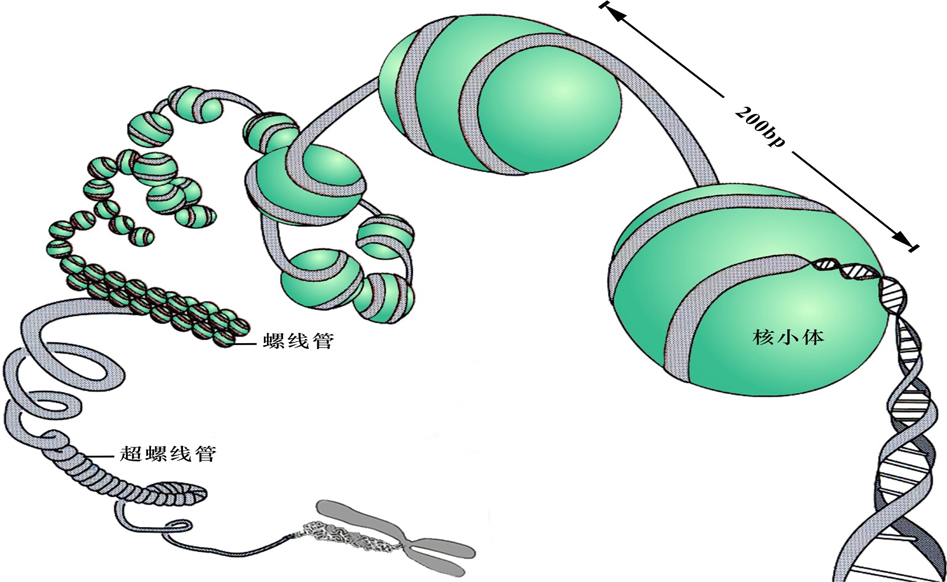
图8-17 染色质组装的多级螺旋化模型
2.染色体骨架-放射环模型(scaffold radial loop structure model)
一级结构——核小体
二级结构——螺线管(30nm染色质纤维)
高级结构:袢环结构、染色单体
螺线管以后的高级结构由30nm染色质纤维折叠成的袢环构成袢环沿染色体纵轴由中央向周围伸出,形成放射环;每18个袢环呈放射状排列形成微带(miniband),微带是染色质高级结构的组成单位;约106个微带沿纵轴排列形成染色单体。
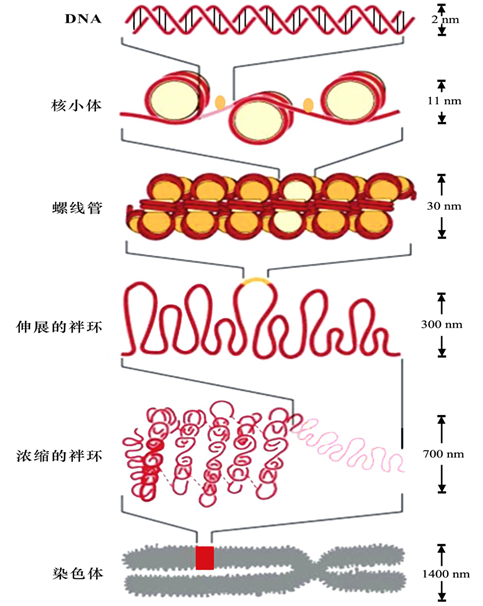
图8-18 染色质组装的放射环模型
视频来源:中国大学慕课网
三、染色质组装形成染色体
20世纪70年代以前,染色质一直被认为是由组蛋白包裹在DNA外,形成类似“铅笔”状的结村 1974年经R.D. Kornberg等人对染色质进行酶切降解研究及电镜观察后,人们对于染色质的结构才有 了进一步的认识。现已知道,染色质的基本结构单位为核小体,核小体在串联的基础上,发生进一步折叠、压缩形成高级结构,最终组装成染色体。
(ー)核小体为染色质的基本结构单位 组成染色质的基本结构单位是核小体(nucleosome)。每个核小体包括有200个左右bp的DMA 个组蛋白分子组成的八聚体及一分子组蛋白H。八聚体是由四种组蛋自H2A、HDB、HB和H各 个分子组成,两个HB、H4二聚体相互结合形成四聚体,位于核心颗粒中央,两个HD2A、HB二聚体 别位于四聚体两侧。146bp的DNA分子在八聚体上缠绕1.75圈,形成核小体的核心颗粒。在两个相邻的核小体之间以连接DNA( linker DNA)分子相连,典型长度约60bp,其长度变异较大,随细胞类型 不同而不同,其上结合一个组蛋白分子H,组蛋白H锁定核小体DNA的进出端,起稳定核小体的作 用。多个核小体形成一条念珠状的纤维,直径约为10mm(图8-10)。
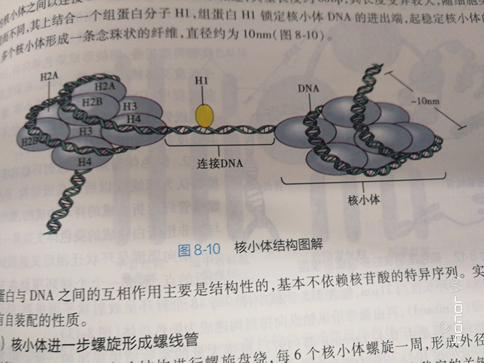
组蛋白与DNA之间的互相作用主要是结构性的,基本不依赖核昔酸的特异序列。实验表明,核 小体具有自装配的性质。
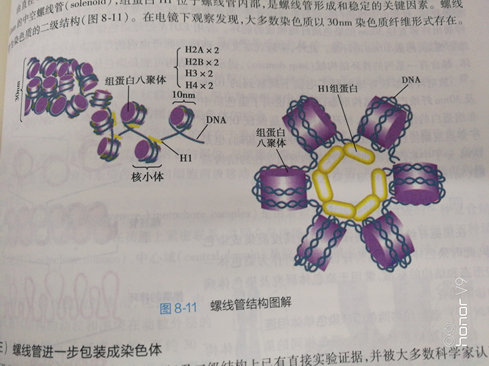
(二)核小体进一步螺旋形成螺线管由直径10nmn的核小体串珠结构进行螺旋盘绕,每6个核小体螺旋一周,形成外径30m,内径 onm的中空螺线管(solenoid),组蛋白H位于螺线管内部,是螺线管形成和稳定的关键因素。螺线 管为染色质的二级结构(图8-11)。在电镜下观察发现,大多数染色质以30m染色质纤维形式存在。
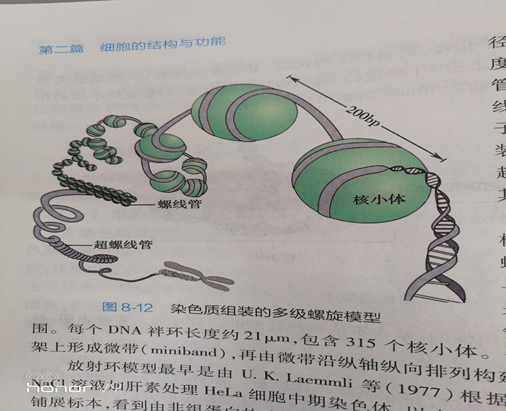
(三)螺线管进一步包装成染色体关于DNA如何组装成染色体,在一级及二级结构上已有直接实验证据,并被大多数科学家认可。从30nm的螺线管如何进一步组装成染色体的过程尚存在争议,目前主要有多级螺旋模型(multiple n2 smodel)及骨架-放射环结构模型( scaffold- radial loop structure model)得到较为广泛的接受。1染色体多级螺旋模型在该模型中,由螺线管进一步螺旋盘绕,形成直径为400mm的圆筒状 构称为超螺线管( supersolenoid),这是染色质组装的三级结构。超螺线管再进一步螺旋、折叠形成美色质的四级结构一一染色单体。根据多级螺旋模型,当DNA分子缠绕在直径10nm的核小体核心颗粒上时,长度被压缩7倍;直径10m的核小体形成螺线管后,DNA分子长度又被压缩6倍;而当螺线管盘绕形成超螺线 管时,DNA分子长度被压缩约为40倍;超螺 线管再度折叠、缠绕形成染色单体后,DNA分 子长度又将被压缩5倍。因此,在染色质的组 装过程中,DNA分子在经过核小体、螺线管 超螺线管到染色单体四级连续螺旋、折叠后,其长度共压缩了8400倍(图8-12)。
2.染色体骨架一一放射环结构模型该模型认为螺线管以后的高级结构,是由30 螺线管纤维折叠成的袢环构成的,螺线管一端 与由非组蛋白构成的染色体支架某一点结合,另一端向周围呈环状迁回后又返回到与其相 邻近的点,形成一个个祥环围绕在支架的周 围。每个DNA祥环长度约2lum,包含315个核小体。每18个袢环呈放射状平面排列,结合在核骨架上形成微带( miniband),再由微带沿纵轴纵向排列构建成为染色单体(图8-13)。放射环模型最早是由U.K. Laemmli等(1977)根据大量的实验结果提出的。他们用2mol/L的 aCl溶液加肝素处理Hela细胞中期染色体,以去除组蛋白及大部分非组蛋白。电镜下观察染色体 甫展标本,看到由非组蛋白构成的染色体骨架,两条染色单体的骨架相连于着丝粒区。由骨架的一点申展出许多直径30mm的染色质纤维构成的侧环。若用EDTA处理染色体标本后,则可见30nm的纤伸展出许多直径30m的染色质纤维构成的侧环。若用EDTA处理染色体标本后,则可见3m 维解螺旋,形成10m的纤维。此外,实验观察发现,两栖类卵母细胞的灯刷染色体和昆虫的多线 体,都含有一系列的祥环结构域( loop domain),提示袢环结构可能是染色体高级结构的普遍特征 放射环模型较好地解释了电镜下观察到的10mm 及30nm纤维产生的结构形态,同时也说明了染色质中 非组蛋白的作用。而且,祥环结构可能是保证DNA分 子多点复制特性的高效性和准确性的结构基础;也是 DNA分子中基因活动的区域性和相对典立性的结构基础(动画8-3“核小体浓缩成染色体的过程”)。